0
发表咨询在线!

发布时间:2020-03-05所属分类:医学论文浏览:1749次
摘 要: 摘要: 超级增强子是由多个相邻近的普通增强子组成的、驱动调控细胞身份基因表达的一个大簇,该区域富集高密度的转录因子、辅因子及增强子相关表观修饰。超级增强子所驱动的异常转录基因对维持肿瘤细胞特性至关重要。肿瘤细胞通过组装自身超级增强子,显著促

摘要: 超级增强子是由多个相邻近的普通增强子组成的、驱动调控细胞身份基因表达的一个大簇,该区域富集高密度的转录因子、辅因子及增强子相关表观修饰。超级增强子所驱动的异常转录基因对维持肿瘤细胞特性至关重要。肿瘤细胞通过组装自身超级增强子,显著促进多种癌基因表达,从而增强肿瘤细胞的增殖、侵袭和转移的能力;抑制超级增强子的活性,则显著抑制肿瘤细胞的生长和存活。本文对目前报道的肿瘤细胞中超级增强子的结构特征和功能调控,以及靶向超级增强子药物研发现状进行了总结,旨在为研发新的针对超级增强子为靶点的抗肿瘤药物提供理论基础和借鉴。
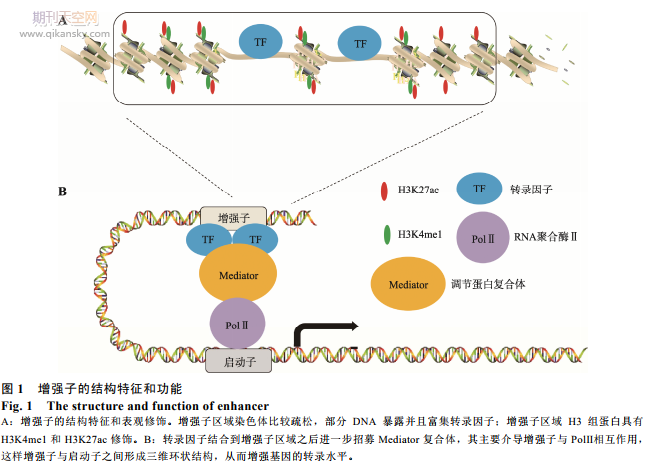
关键词: 增强子;超级增强子;转录;癌症
20 世纪 80 年代,研究发现 SV40 病毒的一段 DNA 序列对于家兔(Oryctolagus cuniculus)的 β-珠蛋白(β-globin,一种能够通过铁卟啉环可逆性结合氧的呼吸性蛋白质)的转录具有增强作用,因此将这一段 DNA 称为增强子(enhancer)[1]。随后的研究发现在哺乳动物细胞内也存在类似特性的 DNA 序列,可以远距离、无方向性的增强基因转录[2~4]。近 30 年研究证明增强子具有以下特征(图 1)[2~5]:(1) 增强子 DNA 序列处于染色体疏松的区域,与核小体中组蛋白的修饰,转录因子的结合有关;(2) 增强子活性与其 DNA 序列结合的组蛋白 H3 的第 4 位赖氨酸单甲基化(H3K4me1)和第 27 位赖氨酸乙酰化(H3K27ac) 修饰程度成正相关[6];(3) 增强子发挥功能需要增强子区域和启动子的区域的直接相互作用,形成三维环状结构(3D-loop)。增强子和启动子的相互作用由多种蛋白介导,如 Mediator 复合体、Cohesin 等[6,7]。
推荐阅读:肿瘤方向文章能投稿哪些学术类期刊
随着 DNA 测序技术的发展,人类对基因有了进一步的认识,对于增强子的研究也越来越深入。2013 年,美国 Young R.A.教授(Whitehead Institute for Biomedical Research)基于当时增强子的研究首次提出超级增强子(super enhancers,SEs)这一概念。他们发现胚胎干细胞(embryonic stem cells,ESC)的主要转录因子结合在一些特殊的增强子上,这些特殊的增强子对于维持胚胎干细胞的干性至关重要,并将这些特殊的增强子定义为超级增强子[8]。超级增强子简单来说就是由多个增强子组成的一个大簇,富集高密度的转录因子、辅因子和增强子表观修饰。它和普通增强子在序列大小、转录因子的结合密度、激活转录的能力以及对转录因子抑制剂的敏感性均不同[8]。随后的研究不仅发现超级增强子存在于多种细胞类型中,也进一步明确了超级增强子区别于普通增强子的功能特性(图 2)[6]:(1) 超级增强子具有高密度的 H3K27ac 和 H3K4me1 修饰,以及 Mediator 复合体和 Bromodomain containing 4 蛋白(BRD4,与组蛋白乙酰化修饰位点结合)的结合;(2) 超级增强子结合的转录因子以及与转录活性相关的染色体的标记比普通增强子高很多;(3) 超级增强子调控的基因比普通增强子调控的基因表达水平高很多;(4) 组成超级增强子的单个增强子也可以像普通增强子一样激活基因转录;(5) 超级增强子可以结合组织中特异的转录因子;(6) 与普通增强子相比,超级增强子活性对于转录因子的阻断更敏感[9,10]。这些现象支持一个假说:超级增强子发挥功能需要结合到超级增强子上的转录因子的合作协同,具有大量转录因子结合的增强子对于基因转录的调控会对转录因子浓度的改变更敏感[11](图 2)。有趣的是,富集在超级增强子上的主要的转录因子也受超级增强子的调控转录,这就意味着超级增强子调控基因转录存在正反馈协同作用,也就形成了细胞中的核心转录调控环路 (core transcription regulatory circuitry, CRC)[12,13]。正是由于超级增强子调控基因表达的特性和其敏感性,因而才能够协调细胞在生长、发育、分化和疾病等各种状态的过渡[9,14~16]。本文主要从超级增强子与肿瘤细胞的关系、在肿瘤细胞中的调控以及该靶点药物在肿瘤治疗中的现状这 3 个方面阐述超级增强子在肿瘤细胞中的作用。
1 超级增强子与肿瘤的关系
2013 年,Young R.A.教授发现在多发性骨髓瘤细胞中超级增强子的区域募集了高浓度的 Mediator 复合体和 BRD4[17],这意味着在多发性骨髓瘤细胞中超级增强子处于活化状态。功能分析实验表明,超级增强子调控的基因(MYC、IRF4、PRDM1、XBP1) 对于多发性骨髓瘤的发生和发展起到关键的促进作用[17]。后续研究发现,超级增强子在多种肿瘤中均有报道,如弥漫性大 B 细胞淋巴瘤[18]、T 细胞急性淋巴细胞白血病[19,20]、默克尔细胞癌[21]、急性髓性白血病[22]、小细胞肺癌[10]、卵巢癌[23]、上皮癌[24]、鳞状细胞癌[25]、黑色素瘤[15]、乳腺癌[26]、食管鳞状细胞癌[27]和结肠癌[28]等。在肿瘤细胞中超级增强子调控的关键癌基因在正常细胞中是不表达的,这就提示超级增强子通过调控这些基因而对肿瘤生成和肿瘤特性维持起到关键作用[9,29,30]。由于细胞在癌变过程中大多数的超级增强子是重新形成具有功能性的元件,因此超级增强子的活化可以作为细胞癌变的一种标志[9,29,30]。综上所述目前研究均表明超级增强子的激活可以促使正常细胞向肿瘤细胞的恶性转化。超级增强子不但对蛋白编码基因具有转录激活作用,对非编码基因,如 microRNA (miRNA,一种长度约 22nt 的小 RNA)的转录及成熟也具有调控功能。美国麻省理工学院生物系 Phillip A. S.教授研究组利用 CRISPR/Cas9 基因组编辑方法发现超级增强子不仅促进 miRNA 的转录,也可以通过招募 Drosha/DGCR8 蛋白复合体促进前体 miRNA (primiRNA)的成熟,以此来调控细胞种类特异性 miRNA 的生成[31]。对 18 种肿瘤细胞分析发现,在有些肿瘤细胞中超级增强子活性上调,而有些肿瘤细胞中超级增强子活性下降。进一步分析表明在细胞癌变过程中激活的超级增强子往往与促癌 miRNA 相关,而失活的超级增强子主要调控抑癌 miRNA 的生成[31]。以上研究提示,调控 miRNA 的超级增强子活性与肿瘤发生发展密切相关。因此,超级增强子联合多个 miRNA (SE-miRNA)将有潜力成为细胞癌变的生物标志物[31],对于肿瘤的早期诊断以及治疗具有重要的临床意义。除此之外,超级增强子还可以调控长链非编码 RNA (long non-coding RNA, lncRNA)的转录[32]。在鳞状细胞癌组织中发现受超级增强子调控的 lncRNA LINC01503 明显上调。进一步研究发现, LINC01503 的表达水平与鳞状细胞癌病人预后呈负相关:LINC01503 高表达的病人生存率低。研究表明激活的增强子或超级增强子区域也可以被转录产生 RNA,称为 enhancer RNA (eRNA),eRNA 可以协同超级增强子激活转录[33,34]。超级增强子发挥功能不仅依赖于和启动子之间的 3D-loop 的形成,也依赖于超级增强子转录的 eRNA 的生成。因此,在临床上可以结合 lncRNA 以及 eRNA 的水平对病人进行精准治疗。
肿瘤的异质性很大一方面是由于一个肿瘤内的细胞通常可能来源于多个不同的细胞克隆,而这些不同克隆来源的肿瘤细胞其超级增强子的激活也存在差异,这就为区分肿瘤亚型或肿瘤细胞亚群提供了一种新的鉴定方法。例如,通过以往的方法对成神经管细胞瘤的生物化学和遗传学分析把其分成 4 个亚型。但是通过对这 4 个亚型的增强子图谱分析发现了一种新的亚型,这种新型的成神经管细胞瘤细胞中都具有与肿瘤异质性相关的超级增强子群[35]。更为重要的是通过分析在这类肿瘤细胞的超级增强子调控的转录因子可以明确细胞特异性的核心转录调控环路(CRC)。通过对于 CRC 分析确定了 LIM homeobox transcription factor 1 alpha (LMX1A,一种转录因子)在第 4 类亚型的成神经管细胞瘤是一个主要转录因子(master transcription factor)[36]。同样,在其他基因异质性癌中也发现类似情况,如三阴性乳腺癌依靠超级增强子调控的特异性的基因群来维持细胞生长和增殖[26]。可见通过对于不同肿瘤细胞的增强子的图谱分析可以独立预测肿瘤亚型,发现之前治疗的不足以及新的潜在治疗靶点,为肿瘤治疗提供新思路、新方向[6]。
综上所述,在多种肿瘤细胞中均发现超级增强子处于异常激活状态,其对于靶基因的调控呈多样化:促进 mRNA 的生成、促进 miRNA 的转录以及成熟、促进 lncRNA 的转录生成以及超级增强子自身转录生成的 eRNA 对于其活性也起到协同作用。除此之外,通过绘制肿瘤细胞的增强子图谱可以预测肿瘤亚型,为基因异质性肿瘤提供统一的治疗平台。
2 肿瘤细胞中超级增强子的调控
在肿瘤细胞中超级增强子的调控是如何实现的呢?早期对于小鼠胚胎干细胞发育的研究提出一个模型:组成超级增强子的每一个增强子都具有活性,而超级增强子的功能类似于一个平台,这个平台汇集了与发育相关的信号通路传递过来的信号,这些信号协同调控超级增强子活性启动基因转录(图 3)[11]。同样,与癌基因相关的超级增强子也富集了肿瘤细胞依赖的信号通路的转录因子。在 Wnt 信号通路异常引起的结肠癌细胞中,相关的超级增强子区域富集了很多由 Wnt 信号通路终端的转录因子 4 (transcription factor 4, TCF4),通过激活或者抑制 Wnt 信号通路,可以控制超级增强子调控的基因转录[11,37]。在雌激素受体(estrogen receptor, ER)阳性的乳腺癌细胞中,相关的超级增强子区域聚集了大量的 ERα;而在三阴性乳腺癌细胞中缺少类固醇激素的表达,与其相关的超级增强子区域富集了完全不同的转录因子[26,37]。
在肿瘤细胞中,信号通路从多方面对超级增强子的活性进行调控。2015 年美国西北大学 Licht J.D. 教授团队研究发现 Ras-Erk 活性与超级增强子的活性密切相关:抑制 Ras 蛋白的活性会导致超级增强子相关的特征(如 H3K27ac)消失、活性下降、降低相关基因转录;激活 Ras 可以增强调控癌基因的超级增强子活性[38]。另一方面,促癌信号通路可以通过操纵转录机器调节超级增强子的活性。转录暂停是激活的 RNA 聚合酶Ⅱ(RNA polymerase Ⅱ,Pol Ⅱ) 在启动子附近停止转录的一种状态[39]。在正常的肝细胞中,Hippo 信号通路可以通过限制暂停的 Pol Ⅱ 释放,因而抑制了增强子或超级增强子调控的基因转录[40]。然而,在肝癌细胞中 Hippo 信号通路的缺失导致 YAP(Yes associated protein)入核,YAP 蛋白结合到超级增强子上,招募 Mediator 复合体和细胞周期素依赖性激酶 9(Cyclin-dependent kinase 9, CDK9),使暂停的 Pol Ⅱ进入到延伸状态,促进癌基因转录[41]。因此,在肝癌中 YAP 通过激活超级增强子促进癌基因的转录。
以上研究表明超级增强子可以作为连接癌基因信号通路和维持肿瘤细胞特性的基因转录表达的渠道。然而进一步研究发现信号通路对于超级增强子的调控与转录因子在超级增强子区动态结合有关。例如,在 NOTCH1 异常导致的 T 细胞白血病(T-ALL) 细胞中,NOTCH1 在基因组上具有普遍的结合,但是只有不到 10%的基因对于 NOTCH1 信号通路的改变有应答,而这些应答基因的 NOTCH1 结合在对应的超级增强子上。如果这些位点丢失 NOTCH1 的结合就会导致超级增强子的特征消失[42]。
声明:①文献来自知网、维普、万方等检索数据库,说明本文献已经发表见刊,恭喜作者.②如果您是作者且不想本平台展示文献信息,可联系学术顾问予以删除.

SCISSCIAHCI

