发布时间:2022-01-19所属分类:农业论文浏览:1次
摘 要: 摘要:解磷微生物能够活化土壤中的难溶性磷,筛选杉木(Cunninghamialanceolata)根际高效解磷菌对于缓解南方红壤区杉木人工林土壤的磷素受限问题具有重要现实意义.以南方红壤区不同林龄(2,4,10,15a)杉木人工林下的根际土壤为研究对象,通过平板分离初筛菌株、液体发
摘要:解磷微生物能够活化土壤中的难溶性磷,筛选杉木(Cunninghamialanceolata)根际高效解磷菌对于缓解南方红壤区杉木人工林土壤的磷素受限问题具有重要现实意义.以南方红壤区不同林龄(2,4,10,15a)杉木人工林下的根际土壤为研究对象,通过平板分离初筛菌株、液体发酵复筛菌株和16SrDNA 测序,筛选、鉴定高效根际解磷菌,采用单因素实验和正交试验确定高效解磷菌的最优培养条件.实验结果表明:1)15a杉木根际土壤的解无机磷菌与解有机磷菌数量分别为3.64×105和2.14×105cfu/g,均显著高于其他林龄(p<0.05),且各林龄解磷菌类型间存在显著差异(p<0.05);2)筛选出25株解无机磷菌和20株解有机磷菌,经平板初筛、液体发酵复筛和16SrDNA 测序鉴定,分别得到一株解磷效果显著(p<0.05)的解无机磷菌株 W1(溶磷量238.08μg/mL,不动杆菌属(Acinetobactersp.))和解有机磷菌株 Y9(溶磷量15.04μg/mL,克雷伯氏菌属(Klebsiellasp.));3)优化后 W1的最佳培养条件为1.0%(质 量 分 数)葡 萄糖、1.50%(质量分数)酵母粉、初始pH7.5、装液量30mL、接 种 量3%(体 积 分 数)、温 度40 ℃,Y9的最佳培养条件为0.5% 葡萄糖、1.25%酵母粉、初始pH6.0、装液量20mL、接种量9%、温度28℃.上述结果可为杉木根际解磷微生物的开发利用提供数据支持.
关键词:杉木;解磷菌;筛选;鉴定;培养条件优化;红壤区;中国南方
磷是植物生长发育、结构组成和生理生化过程的关键元素之一,在农业生产上常通过大量施用磷肥来满足作物对磷元素的需求,但磷肥极易与金属离子形成不溶性磷酸盐进而对磷素起到固定作用[1],这极大限制了肥料的利用效率[2-3].因此,如何活化土壤中难溶态磷并提 高 其 转 化 利 用 效 率 成 为 当 前 土 壤 化 学 的研究热点之一[4].
我国南方林区土壤的有效磷含量极低[5],95%~99%的磷以难溶态存在[6],严重制约着南方重要用材树种杉 木(Cunninghamialanceolata)人 工 林 的 可 持续经营[7-9].针对杉木人工林经营中面临的低磷胁迫,诸多学者从施肥[10]、树种共生[11]、硅肥配施[12]等方面做了 大 量 有 益 尝 试,虽 取 得 了 一 些 成 效,但 依 旧 无 法有效、低成本地改善杉木人工林下土壤速效磷短缺的情况.针对南方林区杉木林地的磷素受限问题,如 何有效提升杉木对磷的利用效率,对杉木人工林的可持续经营具有重要的现实意义.
活跃在植物-土壤接触面上的微生物群落对于维持植物生长发育和植物健康起着重要作用[13].现有研究认为根际微生物是农作物根际的核心,在改善土壤理化性质方面发挥了极大的作用[14-16].利用根际微生物改善林木对营养元素的吸收状况已有诸多尝试,前期研究从 红 树 林(mangrove)[17]、巨 尾 桉(Eucalyptusgrandis)[18]、马 尾 松 (Pinusmassoniana)[19]、枫 香(Liquidambarformosana)[20]、降 香 黄 檀 (Dalbergiaodorifera)[21]等植 物 根 际 筛 选 出 了 一 批 具 有 显 著 解磷效果的根际微生物[22-23],这为提升杉木在困难立地条件下的养分吸收效率提供了新思路.吴则焰等[24]提出杉木连栽导致土壤养分逐代降低,进而表现为不同林龄杉木的根际微生物群落存在较大差异.杉木根际磷素含量随着林龄的增加而降低[25],中幼龄杉木人工林下的土壤微生物数量显著高于其他林龄[26],且中幼林对 养 分 需 求 量 大.因 此,本研究以不同林龄杉木 人工林 为 研 究 对 象,以 杉 木 根 际 土 壤 为 供 试 土 壤,通 过平板定性与摇瓶定量筛选出高效解磷菌,鉴定高效解磷菌 株 类 型.在 此 基 础 上,通过单因素实验与正交 试验优化高效解磷菌生长条件,以期获得具有较高解磷能力和生长量的菌株,为利用根际微生物提升杉木人工林对根际无效磷的吸收利用效率奠定实验基础,亦可探索利用 解 磷 菌 改 善 杉 木 人 工 林 地 力 衰 退 问 题 的可行性.
1 材料与方法
1.1 实验材料
供试土壤样品采集于福建省南平市建阳区溪东国有林场(118°08′~120°31′E,26°40′~27°20′N),地处武 夷 山 脉 南 侧,属 于 典 型 的 中 亚 热 带 季 风 气 候,冬温夏热,四季分明,季风发达,年均降水量1700mm,年蒸发量1500mm,年均温大于18℃,十分有利于杉木的 生 长 发 育.林 场 前 身 为 低 产 低 效 马 尾 松 人 工 林,2008—2010年逐年砍伐后营造杉桐混交林[27],林 下植被 种 类 主 要 有 苦 竹 (Pleioblastusamarus)、芒 萁(Dicranopterisdichotoma)、观 音 座 莲(Strobilanthescyclus)、黄瑞木(Adinandramillettii)等.
1.1.1 土壤样品采集及理化性质测定
本实验选取中幼林龄(2,4,10,15a)杉木人工林,各取3块具有代表 性 的20m×20m 样 地,每 块 样 地采用五点取样法,铲去表土后深挖10~20cm,选取具有完 整 根 系 的 土 体,采用抖落法采集根际土壤,同 一样地各取样点土壤均匀混合并标号处理,同时采集非根际土壤进行标号处理,将土样装入冰盒带回至冰箱(4 ℃)保存.其中新鲜根际土样用于菌株筛选,非根际土样待风干过筛后进行理化性质测 定(表1),土 壤 类型均 为 红 壤,其他理化性质采取常规测定方式:全 磷测定采用钼锑抗比色法,有效磷测定采用盐酸-硫酸浸提法,有机质测定采用重铬酸 钾-外 加 热 法,全 氮 测 定采用半微量凯氏法,水解氮测 定 采 用 碱 解-扩 散 法,全钾测定采用碱熔-火焰光度法,速效钾测定采用乙酸铵浸提-火焰光度法,pH 测定采用电位法[28].
1.1.2 培养基
李豆豆等[29]指出以磷酸三钙为磷源时,菌株解磷量显著高于其他磷源类型,因此采用磷酸三钙无机磷培养基(葡萄糖10.0g,硫酸铵0.5g,硫酸镁0.3g,氯化钠0.3g,氯化钾0.3g,硫酸亚铁0.03g,硫酸锰0.03g,磷酸三钙5.0g,琼脂18.0g,蒸馏水1L,pH7.0~7.5)来 分 离 解 无 机 磷 菌 株.采用蒙金娜有机磷培养基(葡萄糖10.0g,硫酸铵0.5g,硫酸镁0.3g,氯化钠0.3g,氯化钾0.3g,硫酸亚铁0.03g,硫酸锰0.03g,卵磷脂0.2g,碳酸钙5.0g,琼脂18.0g,蒸馏水1L,pH7.0~7.5)来分离解有机磷菌株[30].发 酵基础培养基选用 LB液体培养基(胰蛋白胨10.0g,酵母浸出粉5.0g,氯化钠0.5g,pH7.0~7.2)[31].
1.2 解磷菌的筛选、鉴定及培养条件优化
1.2.1 解磷菌分离与纯化
取摇床震 荡 后 的 土 壤 溶 液 上 清 液 成 倍 数(10-3,10-4,10-5)稀释,涂于磷酸三钙无机磷培养基和蒙金娜有机 磷 平 板 培 养 基 上,每 个 梯 度 设 置3组 重 复,置于28 ℃恒温培养 箱 中(解 有 机 磷 菌 培 养3d,解 无 机磷菌培养7d)[32-33],记 录 含 溶 磷 圈 菌 落 数.菌 落 的 纯化采用划线法并培养3~7d,纯化后将单菌落转移至牛 肉 膏 蛋 白 胨 斜 面 培 养 基 上,保 存 于 4 ℃ 冰 箱备用[34].
1.2.2 解磷菌筛选
解磷菌筛选采用平板初筛与摇瓶复筛.平板初筛需记录各菌株的溶磷圈直径、菌 落 直 径 及 二 者 比 值,各菌株 设5组 重 复,取 平 均 值.摇 瓶 复 筛 需 将 初 筛 得到的菌株接种到液体培养基中,摇床培养7d(28 ℃,160r/min),对 培 养 好 的 菌 株 进 行 离 心 处 理 (4 ℃,10000r/min,10 min),利用钼锑抗比色法检测上清液中的溶磷量,判断各菌株的溶磷能力.
1.2.3 16SrDNA 测序鉴定
利用16SrDNA 通用引物序列 F27和 R1492对筛选得到的细菌进行扩增,纯化后送往上海迈浦生物科技有限公司进 行测序,将 测 序 结 果 提 交 至 RDP(http:∥rdp.cme.msu.edu/)及 NCBI(http:∥www.ncbi.nlm.nih.gov/)数 据 库 中 进 行 序 列 比 对 分 析,选取与 GenBank中同源性最高的序列,初步鉴定菌株.
1.2.4 培养优化
培养基组分 优 化:采 用 不 同 碳 源(葡 萄 糖、蔗 糖、乳糖、可 溶 性 淀 粉、麦 芽 糖、甘 露 醇)代 替 基 础 培 养 基中的碳源,根据菌液在600nm 下的吸光度(A600)确定最佳 碳 源.改 变 最 佳 碳 源 质 量 分 数 (0.5%,1.0%,1.5%)进行菌株培养以确定最佳浓度.最 佳 氮 源(硫酸铵、氯 化 铵、硝 酸 钾、蛋 白 胨、酵 母 粉、尿 素)及 其 最适质量分数(0.5%,0.8%,1.0%,1.3%,1.5%)的确定采取相同方式.
培养条件优 化:在其他条件不变的情况,分 别 改变菌株 发 酵 液 的 pH(5.0,6.0,6.5,7.0,7.2,7.5,8.0,9.0)、装液量(10,20,30,40,60mL,于100mL发酵瓶)、接 种 量 (1%,3%,5%,7%,9%,均 为 体 积 分数)、培养温度(20,25,28,30,35,40 ℃),测 量 对 应 的A600,确定对应的最适值.
正交试验:根 据 单 因 素 实 验 结 果,设 计 四 因 素 三水平正交试验,基于初始pH 值(A)、装液量(B)、接种量(C)、培养温度(D)以及不同水平的培养条件进行优化,每个处理设3个重复.
1.3 数据处理
采用 Exce12010软件进行原始数据的整理、分析及图像绘制,运用SPSS19.0软件进行单因素方差分析(显著水平0.05)和 最 小 显 著 差 数(LSD)法 多 重 比较,采用 Pearson相关系数法确定 pH 与 溶 磷 量 之 间的相关关系,运 用 正 交 设 计 助 手Ⅱ v3.1处 理 正 交 试验数据.
2 结果与分析
2.1 解磷菌的筛选结果
对分离出的菌落进行形态特征判断,经福建省林业科学研究院形态学鉴定分析其培养形态,发现4种不同林龄的杉木根际土壤解磷菌形态各异.所筛选出的解磷菌以不透明的白色、乳白色、浅黄色为主,菌落呈圆形、不规 则 形,边 缘 基 本 整 齐,中 间 以 凸 起 为 主,菌株 表 面 大 多 光 滑.不同菌株的生长速度不一致,绝大多数菌株的生长速度较快,可在24h内生长为菌落成型;但亦存在解磷真菌在48h后才长势较好,生长速度较缓慢.
不同林龄杉木根际解磷菌数量及种类亦存在差异(表2).从数量来看,杉木人工林下根际解无机磷菌的数量范围为2.12×105~3.64×105cfu/g(cfu为菌落形成单位),解有机磷菌的数量范围为1.31×105~2.14×105cfu/g.其中,15a杉木根际土壤的解无机磷菌数量和解有机磷菌数量均显著高于其他林龄(p<0.05),且 类 型 数 最 多,而解有机磷菌类型数为 9,与4a杉木的类型数相同.
从以上 解 磷 菌 中 挑 选 解 磷 效 果 具 有 显 著 优 势 的菌株进行溶磷实验,对高效解磷菌进行16SrDNA 测序鉴 定.通 过 对 杉 木 根 际 解 磷 菌 进 行 平 板 初 筛,无 机磷培养基和有机磷培养基中均出现较明显的透明圈,表明根际解 磷 微 生 物 能 够 溶 解 无 机 磷 和 有 机 磷 并 转化至可被植物或自身吸收利用的有效磷素.本研究共筛选得到具有较明显作用的25株解无机磷菌株和20株解 有 机 磷 菌 株.对 以 上 菌 株 进 行 摇 瓶 复 筛 后 发 现,在固体培养 基 中 溶 磷 圈 直 径 与 菌 落 直 径 比 值 大 的 菌株,在液体培养基中的解磷能力并不一定显著突出,这与黄鹏飞等[35]关于解磷菌在固体平板和液体培养两种方式下 的 解 磷 效 果 并 不 存 在 线 性 关 系 的 研 究 结果一致.图中数据字母不同表示差异显著(p<0.05),下同.图2 解无机磷菌株的溶磷量Fig.2 Phosphorussolubilizingcapacityofinorganicphosphorus-solubilizingstrain
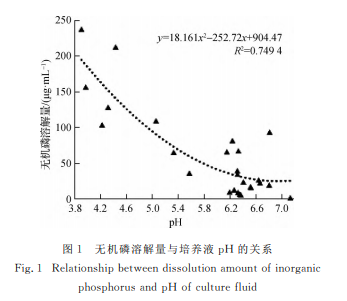
解磷菌常通过分泌有机酸来溶解难溶性磷,发酵液的pH 降 低,从 侧 面 可 反 映 出 菌 株 溶 磷 能 力 的 高低[36-37].本研究 在 探 究 解 无 机 磷 菌 株 的 溶 磷 能 力 时,对菌株培养液的pH 进行测定,所测值均低于对照组,这一结果进一步验证了上述结论.发酵液 的无机磷溶解量与pH 呈极显著负相关性(p<0.01,R2=0.7497,r=-0.8659),解磷能力强的菌株培养 液 pH 较 低,反之pH 较高,只有个别菌株呈现差异(图1).有机磷溶解量 与 pH 的 相 关 关 系 并 不 显 著(p>0.05),解 有机磷菌株的溶液pH 主要集中于6.58~7.42之间,仅菌株 Y3溶液的pH 为1.89,呈现出强酸性,与其他菌株差异较大,究其原因,可能是由于大部分解磷菌以酶解为主[38],而 Y3以分泌有机酸来发挥溶磷作用[39-40].
通过复筛,采用单因素方差分析各菌株的无机磷溶解量.无 机 磷 溶 解 量 较 空 白 对 照 组(CK)均 有 增 加(图2),增量范围为4.01~235.88μg/mL,溶 磷 量 范围为6.31~238.08μg/mL.W1的解磷效果显著高于其他菌株(p<0.05),溶磷量达238.08μg/mL;溶 解无机磷效果最差的是 W15,溶磷量仅为6.31μg/mL.W1溶磷量是 W15溶磷量的37.73倍.
有机磷溶解量较空白对照组均有增加(图3),增量范围为1.07~11.33μg/mL,溶磷量范围为4.78~15.04μg/mL.Y9的解磷效果显著高于其他菌株(p<0.05),溶 磷 量 达 15.04μg/mL;解磷效果最差的是Y10,溶磷量 仅 为4.78μg/mL.Y9溶 磷 量 为 Y10溶磷量的3.15倍.
2.2 菌株鉴定结果
经 PCR 扩 增 后 的 16SrDNA 基 因 序 列 (附 录(http:∥jxmu.xmu.edu.cn/upload/html/20220115.html)图S1和 S2)通 过 核 酸 BLAST 序 列 比 对 后,得到相似菌株的登录号与相似度.经鉴定,W1为不动杆菌属(Acinetobactersp.,登录号 MZ145066.1)(图4),Y9为克雷伯氏菌属(Klebsiellasp.,登录号 MZ145064.1)(图5).
2.3 解磷菌培养条件优化
综合上 述 实 验 结 果,选 用 解 无 机 磷 效 果 最 好 的W1与解有机磷效果最好的 Y9进行培养基组分优化和培养条件优化.图5 菌株 Y9的系统发育树Fig.5 PhylogenetictreeofstrainY9
2.3.1 单因素实验筛选
菌株生长量对于不同碳源、氮 源、初 始 pH、装 液量、接种量、温度的响应有显著差异(图6).碳源是微生物进行新陈代谢等活动的主要能量来源[41].6种不 同 碳 源(葡 萄 糖、蔗 糖、麦 芽 糖、乳 糖、甘露醇和可溶 性 淀 粉)对 W1和 Y9的 生 长 量 的 影 响差异显著,W1和 Y9以葡萄糖为碳源时的A600值均显著高于其他碳源时,菌株生长效果最好(图6(a)),其次为甘露醇、麦芽糖、乳糖、可溶性淀粉、蔗糖.故改变最适碳源葡萄糖的质量分 数(图6(b)),W1在1.0%时生长量显著大于其他质量分数时,而 Y9在0.5%时生长量最大.
氮源也是微生物生长发育的重要元素之一.当以酵母粉为唯一氮源时,W1和 Y9的生长量显著大于其他氮源时(图6(c)),其 次 为 蛋 白 胨,且 酵 母 粉 和 蛋 白胨对菌株生长量的影响显著大于其他4种氮源.故设置含不同质量分数酵母粉的培养基,结果表明 W1在1.50% 时生长量显著大于其他处理,而 Y9在1.25%时生长量最 大(图 6(d)),且 质 量 分 数 升 高 时 菌 株 的A600值变化差异 并 不 明 显,表 明 该 菌 株 对 高 浓 度 酵 母粉氮源变化并不敏感.
pH8.0 处 理 下 W1 生 长 量 最 大,但 pH 7.0 和7.5处理下差异 性 并 不 显 著,故后续正交试验采取上述3种初始pH 处理;而 Y9生长量主要在pH5.0~6.0之间呈上 升 趋 势,后 续 A600值 随 着 pH 增 加 而 降低(图6(e)).不 同 装 液 量 下,菌 株 生 长 量 显 著 不 同,W1与 Y9的最适装液量均为30mL(图6(f)).不同接种量下,3%,5%,7%接种量的 W1菌液A600值差异不显著,而 Y9在9%接 种 量 时 生 长 量 显 著 大 于 其 他 处理.温度过高或过低也不利于菌株的生 长,35 ℃处 理下 W1生长量显著大于其他处理,而 Y9生长量在28,30和35 ℃处理下的差异不显著,因此设计正 交 试 验进一步确定最适温度.
2.3.2 正交试验
设计正交试验,选择4个因素(初始 pH 值、装液量、接种量和温 度)及 每 个 因 素 设 定3个 水 平.W1的因素 A 为初始pH 值(7.0,7.5和8.0),因素 B 为装液量(20,30和40mL),因素 C 为接种量(3%,5%和7%)、因素 D 为 温 度(30,35和40 ℃).其 中,因 素 A对 W1生长量影响最大,因素C和 D次之,W1的最佳培养条件为A2B2C1D3,即初始pH7.5、装液量30mL、接种量3%、温度40 ℃(表3).
Y9的因素 A 为初始pH 值(5.0,6.0和6.5),因素 B为装 液 量 (20,30 和 40 mL),因 素 C 为 接 种 量(5%,7%和9%)、因素 D 为温度(28,30和35 ℃).由于RD>RB>RC >RA,4个因素对菌株生长的影响程度为温度>装液量>接种量>初始pH,则 Y9的最佳培 养 条 件 为 A2B1C3D1,即 初 始 pH 为6.0、装 液 量20mL、接种量9%、温度28 ℃(表4).
正交试验结果与单因素实验结果存在差异,表明各因素之间存在交互作用,而正交试验结果更具备准确性.
3 讨论与结论
逆境情况下,植物受环境影响会激发自身的应激性.邹显花等[42]提出杉木根系通过大量增生来应对低磷胁 迫,加快向地生长以应对高磷环境.当 土 壤 有 效磷含量丰富时,林木通过自身调节便可获取足够的营养元素,但处于低磷胁迫条件下的林木除自身的应激性外,还需要解磷微生物来推动土壤中无效磷元素的转化以保证供给.
南方地区杉木人工林下土壤有效磷在固定作用的影响下含量降低,根际解磷微生物的应用能够有效缓解低磷压力[43].本研究在杉木根际筛选出的解无机磷菌 W1的 溶 磷 量 高 达238.08μg/mL,解 有 机 磷 菌Y9的溶磷量达15.04μg/mL.Y9菌株的有机磷溶解量大于范丙全等[44]在杉木根际筛选出的乌博内氏伯克霍尔德菌(Burkholderiaubonensis)的解磷菌P5(溶磷量为195.61mg/L),这可能是因为不同种类的微生物代谢机制具有多样性,进而导致分泌物种类和数量的不同,影响解 磷 微 生 物 的 解 磷 能 力[45];但 也 有 研 究指出,微生物、土壤的空间异质性[46]以及生存环境[47]的差异均会对微生物与植物互作产生影响.
对两株菌株基因序列进行 Blast对比得到 W1为不动杆菌属,Y9为克雷伯氏菌属.李文等[48]提出不动杆菌可以在农业中应用以提高磷素的溶解量,他所筛选出的JL-1菌株溶磷量为118.04mg/L,解磷效果显著弱于 W1.李梦娇等[49]研 究 得 出 克 雷 伯 氏 菌 属(K. pneumoniae)对 于 增 强 植 物 的 溶 磷 能 力 具 有 重 要 作用,本研究的结果验证了高效解磷菌 的溶磷作用显著.庄馥璐等[46]将筛选出的不动杆菌属 PsbM8菌 株回接拟南芥(Arabidopsisthaliana)后,根系附近有明显的溶磷圈出现,进一步得出不动杆菌属在解磷微生物的筛选与鉴定研究中应用较广泛的结论.目前常见报道的解磷菌主要有固氮菌属(Azotobacter)、假单胞菌属(Pseudomonas)、芽 孢 杆 菌 属(Bacillus)、欧 文 氏菌属(Erwinia)、根瘤菌属(Bradyrhizobium)、青霉属(Penicillium)、根 霉 属 (Rhizopus)和 链 霉 菌 属(Streptomyces)[9,34,46].下一步亦可针对不同种属解磷菌的解磷效果差异进行探索.
选用高效菌株进行培养基组分和培养条件优化,明晰菌株的最适宜生存环 境,促进其发挥最大功效,为日后田间根际大规模应用奠定基础.培养基优化实验结 果 表 明:W1 菌 株 与 Y9 菌 株 分 别 以 1.0% 和0.5% 的葡萄糖为最佳碳源,这与李文等[50]提出的不动杆菌的最佳碳源为葡萄糖的结论一致;氮源的最佳选择分别为1.50%和1.25%的酵母粉.南方红壤区多呈酸性,Y9菌株偏向酸性环境,适宜应用于南方杉木根际;而 W1菌株偏向碱性环境,可在实际应用过程中加以调节土壤酸碱度,为 其 生 长 创 造 最 适 环 境.韦 宜慧等[51]在杉木根际筛选出乌博内氏伯克霍尔德菌的P5菌株生长的pH 最适范围为5~6,最适温度为25~30℃.目前发现的解磷菌的最优培养条件偏向高温环境,解 磷 菌 的 功 效 可 能 与 温 度 密 切 相 关,针 对 杉 木 主产区土温较低的情况,有效降低目前现有优势解磷菌的最优培养温度是值得探索的方向.
随着微生物与植物互作机制研究的日趋完善,根际微生物在 解 决 林 木 根 际 营 养 元 素 胁 迫 问 题 领 域 中的应用前景将更加广阔.现有研究主要探讨解磷菌的筛选、鉴定、培养条件优化及其作用机制,以期获得菌株最 大 生 长 量,发挥高效解磷菌的最大效用.王 志 康等[52]提出土壤有机质含量、碳氮比等是解磷微生物发挥作用的限制因子.为了更好地将解磷微生物应用于实际生产,在下一步研究中应深入探讨解磷菌与植物的互 作 机 制,可通过适当调控环境因子,提 升 为 解 磷微生 物 生 存、生长所能提供的最大资源供应量,以 满足接种解磷菌的繁殖与生长需求,从而为利用微生物改善低磷胁迫环境提供最优的菌种资源支持.——论文作者:赵 君1,饶惠玲1,王耘籽1,黄 伟1,吴承祯2,李 键1*
参考文献:
[1] 梁翠月,廖红.植物根系响应低磷胁迫的机理研究[J].生命科学,2015,27(3):389-397.
[2] 黄达明,李倩,管 国 强,等.一株解磷细菌的筛选、鉴 定 及其溶磷培养条件的优化[J].生物技术 通 报,2015,31(2):173-178.
[3] 刘聪,林维,孙珑,等.黑土区林地土壤高效解磷细菌的分离、筛选及其解 磷 效 果[J].东北林业大学学报,2013,41(11):83-85,122.
[4] 高桂凤,党博,蔡柯,等.1株解磷菌株鉴定及影响其解磷能力因素[J].东北 林 业 大 学 学 报,2020,48(1):102-104,109.
[5] 鲁艳红,廖育林,聂军,等.长期施肥红壤性水稻土磷素演变特征及对 磷 盈 亏 的 响 应 [J].土 壤 学 报,2017,54(6):1471-1485.
[6] 詹书侠,陈伏生,胡小飞,等.中亚热带丘陵红壤区森林演替典型阶段土壤氮磷有效性[J].生态 学 报,2009,29(9):4673-4680.
[7] 李冠军,梁安洁,洪滔,等.内生真菌对杉木凋落叶质量损失和养分含量的影响[J].应用与环境 生 物 学 报,2018,24(6):1211-1220.
[8] 艾如波,徐彩瑶,吴承祯,等.低磷胁迫下内生真菌对宿主杉木幼苗生 理 生 化 的 影 响 [J].贵 州 农 业 科 学,2020,48(1):108-113.
[9] 罗扬,刘书影,周柳婷,等.连栽杉木根际土壤镰刀菌属真菌 群 落 变 化 规 律 [J].生 态 学 杂 志,2020,39(9):2921-2922.
[10] 李慧敏,王瑞,施 卫 明,等.菜地土壤解磷微生物特征及其在磷形态转化 调 控 中 的 作 用[J].土 壤,2020,52(4):668-675.
[11] 黄承标,曹继钊,吴 庆 标,等.秃杉林与杉木连栽林的土壤理化性质及 林 木 生 长 量 比 较[J].林 业 科 学,2010,46(4):1-7.
[12] 朱从桦,张鸿,袁 继 超,等.低磷胁迫下加硅对玉米苗期硅、磷营养及叶绿素荧光参数的影响 [J].水 土 保 持 学报,2017,31(1):303-309.
SCISSCIAHCI