0
发表咨询在线!

发布时间:2022-01-22所属分类:农业论文浏览:1924次
摘 要: 摘 要 深入了解荒漠短命植物的化学计量特征,有助于更好地理解生境土壤因子与植物生存策略的关系。以古尔班通古特沙漠广泛分布的尖喙牻牛儿苗(Erodium oxyrrhynchum)、假狼紫草(Nonea caspica)、琉苞菊(Hya lea pulchella)、飘带果(Lactuca undulata)为植物材料,测定

摘 要 深入了解荒漠短命植物的化学计量特征,有助于更好地理解生境土壤因子与植物生存策略的关系。以古尔班通古特沙漠广泛分布的尖喙牻牛儿苗(Erodium oxyrrhynchum)、假狼紫草(Nonea caspica)、琉苞菊(Hya⁃ lea pulchella)、飘带果(Lactuca undulata)为植物材料,测定不同深度(0~5、5~10、10~15 cm)的土壤理化性质,野外原位多时段采样比较分析4种植物化学计量特征与土壤因子的动态变化及其耦合关系。结果表明:①4种短命植物碳(C)、氮(N)、磷(P)含量普遍较低,化学计量特征存在物种间差异,但在不同生长期内变化规律大致相似。N、P含量从幼苗期到结实期逐渐减少,而C含量则长期趋于稳定。整个生长期内,P、C∶P的变异幅度较大,相反,C、N∶P的变异幅度较小。4种植物在不同生长期的化学计量特征差异与生长期、植物种类存在显著相关性。②0~5 cm土层有机碳(SOC)、总氮(TN)、总磷(TP)含量最高,且随土层的加深逐渐减少。随着植物的生长, 0~5 cm土层中SOC和TN含量呈明显增加的趋势,而TP含量却呈现抛物线状变化趋势。在植物不同生长期内,土壤TP含量稳定性最强,SOC、TN的变异性较强,较低的N含量及TN∶TP比显示出该区域土壤属于N素缺乏类型。3个土层土壤在不同生长期的化学计量特征差异受土层、生长期的影响显著。③4种植物与各层次土壤化学计量特征的相关性无一致规律。植物化学计量指标仅与0~10 cm土层SOC、SOC∶TP相关性较强,而大部分化学计量特征间未显示出相关性。上述结果说明植物化学计量特征并非全部由土壤养分特征直接决定,其明显的种间差异显示了植物自身遗传特性在土壤—植物计量特征耦合关系的重要性。
关键词 生态化学计量学;短命植物;荒漠土壤;相关性;古尔班通古特沙漠
生态化学计量学是研究生态系统中多重化学元素平衡的科学,广泛被应用于受限元素判断,物种个体生长,群落动态演替和生态系统稳定等研究领域[1~2]。生态化学计量学可以通过分析生态系统组成部分的元素含量与比值,认识元素耦合关系、共变规律和驱动机制等问题,有助于解决植物对于外界养分供需平衡、适应反馈能力等方面的难题。碳(C)、氮(N)、磷(P)等化学元素不仅是植物生长的必需元素,也是衡量土壤养分肥力的关键指标[3]。土壤养分与植物化学计量特征密切相关,不仅影响植物生长发育,更直接影响到生态系统的物种组成和生产力[4]。因此,研究植物营养元素含量特征,并结合当地土壤化学计量特征,可以深入了解植物生长过程中对于营养元素的吸收利用情况及其生境土壤养分元素的循环状况,阐明植物与土壤之间相互平衡、相互制约的关系。
古尔班通古特沙漠是温带荒漠生态系统,降水稀少、土壤贫瘠,草本植物种类丰富,多为短命植物和一年生草本植物,其中短命植物是一类利用早春雨雪,在夏季干旱到来前快速完成整个生长发育过程的特殊草本植物类群[5]。一般情况下,随着植物的生长,细胞内贮藏性物质的比例会逐渐升高,但正是由于短命植物体内营养元素的非平衡分配才使其能够利用有限的资源快速完成生长繁殖[6]。动态平衡理论认为[7],有机体能够控制自身的多种特性,包括元素含量和 pH 值等,从而保持相对稳定的内部环境,不随外部环境的变化而剧烈变化。但在不同生长季节,土壤 C、N、P 等元素矿化速率不同,加之复杂变化的外部环境,高等植物可能改变其生长模式,不断调整体内营养元素的分配,造成体内元素含量在各生长阶段具有较大的变异性。因此,本研究旨在探讨植物在不同生长发育阶段,其体内营养元素含量是否会产生变化,土壤因子对植物的生长发育是否存在影响,以及整个生长期植物—土壤化学计量特征的相关生长关系。
目前,国内外的生态化学计量学发展迅速,研究领域主要集中在森林、草地、湿地等生态系统[8],然而对于荒漠区植物与土壤化学计量特征的关注相对较少。为此,本研究在古尔班通古特沙漠展开,该沙漠丘间低地多被生物结皮覆盖,但坡上却鲜见生物结皮,土壤存在明显的空间异质性[9],所以选择坡上广泛分布的4种短命植物:尖喙牻牛儿苗(Erodium oxyrrhynchum)、假狼紫草(Nonea caspi⁃ca)、琉苞菊(Hyalea pulchella)和飘带果(Lactuca undulata)与 3 个不同深度(0~5、5~10、10~15 cm)的土壤作为研究对象,比较分析不同生长期植物及其土壤生态化学计量特征,探讨植物及其土壤生态化学计量比随生长期的变化规律,以及土壤化学元素计量比与土壤间的相关关系。该研究有助于进一步诠释古尔班通古特沙漠坡上短命植物与土壤养分元素的循环、平衡及限制状况,深入认识该区域短命植物功能特征和生存策略,可为该沙漠植被的保护与恢复提供科学依据。
1 材料与方法
1. 1 研究区概况
古尔班通古特沙漠位于新疆天山北部的中央,沙漠地跨 44°15′~46°50′N,84°50′~91°20′E,总面积约 4.88 万 km2 ,是我国面积第二大沙漠,也是我国面积最大的固定、半固定沙漠[10]。该地区为典型的内陆温带干旱性气候,夏季炎热、冬季寒冷,年均降水量小于 150 mm,年均蒸发量大于 2 000 mm[11]。土壤组成以细沙为主,含有少量的粉沙和粘土,总体养分含量偏低,多聚积于表层[12]。土壤 pH 碱性至强碱性,成土过程相对缓慢,土壤养分十分匮乏[13]。草本植物种类繁多、数量丰富,多为一年生草本植物和短命植物,主要优势 植 物[14]有 白 梭 梭(Haloxylon Persicum)、梭 梭(Haloxylon ammodendron)、蛇 麻 黄(Ephedra dis⁃ tachya)、淡枝沙拐枣(Calligonum leucocladum)、尖喙 牻 牛 儿 苗(Erodium oxyrrhynchum)、假 狼 紫 草(Nonea caspica)、碱蓬(Suaeda glauca)等。沙漠丘间低地土壤表面广泛分布着生物结皮,是除草本植物外固定沙面的重要生物因子[9]。
1. 2 样品的采集与处理
经前期野外调查,在古尔班通古特沙漠南部(44.37°N,86.12°E),选择一处地势相对较高且有4 种目标植物分布的区域作为大型样方,再将其划分为 6 块(10 m×10 m)的小型样方,每个小型样方之间至少间隔20 m。因物种生长期的差异且沙丘空间异质性较大,在小型样方内又划分出 10 个(2 m×2 m)的小样方,共计60个小样方。根据短命植物生长周期的特点(新疆短命植物一般在3月下旬至4月上旬开始萌发,随后进入20 d左右的快速营养生长期,花期主要在 4 月中旬至 5 月下旬,果实在5月中旬至6月中旬成熟,随后植株地上部分开始干枯或整株死亡,以地下器官或种子渡过休眠期[15])并结合实际观测情况,在植物第二对真叶出现时开始首次取样(2018年4月8日),之后的采样时间分别是4月22日,5月5日和5月20日,这4 个采样时间代表短命植物的不同生长阶段(幼苗期、展叶期、开花期、结实期)。每次进行植物样品采集时,在相距较远的几个小样方内选择长势良好的植株,采用完全挖掘法获取植株,根系尽可能深挖(幼苗期至结实期逐渐加深),保证其完整。每次每种植物共采集30株。采集后用去离子水将植株清洗干净、自然晾干,于烘箱 105 ℃条件下进行15 min杀青,随后在70 ℃条件下烘干至恒质量,最后使用球磨仪研成粉末,装袋待测。
该区域土壤为风沙土,依据土壤空间异质性的差异以及植物根系分布的层次,在每次采集过样品植物的周围,用四分法[16]对土壤进行3个不同深度(0~5、5~10 和 10~15 cm)的采集。土壤样品采样时,要仔细清除土壤表面植物凋落物与生物结皮等杂质,取样后于阴凉处自然风干,过 100目漏筛除去杂质,用精度为 0.000 1 g 的天平称取 100 g土样装袋待测。
1. 3 营养元素的测定
植物与土壤的C含量用重铬酸钾法测定;N含量用凯氏定氮法测定;P 含量用钼锑抗比色法测定[17]。营养元素含量测定前,需再次将样品烘干以防止空气中水分与元素的污染。
1. 4 数据分析
首先对4种短命植物和土壤不同生长期的C、 N、P含量,C∶N、C∶P和 N∶P进行正态性检验,再利用 Levene’s test 检验方差齐性。并对以上指标进行单因素方差分析(One-way ANOVA)和 Duncan、 T2 Tamhane’s test多重比较,采用双因素方差分析分别对不同生长期、不同植物的 C、N、P 含量, C∶N、C∶P和N∶P,不同生长期、不同土层土壤的C、 N、P含量,C∶N、C∶P和 N∶P进行差异性检验,其中极显著性差异为 P<0.01,显著性差异为 P<0.05。采用变异系数法(Coefficient of variation),比较整个取样时期 4种短命植物和土壤化学计量特征的动态变化(强变异:CV≥0.35;中等变异:0.15≤CV< 0.35;弱变异:0≤CV<0.15)[18]。采用Pearson相关分析法分析植物与土壤化学计量特征之间的相关性。常规数据的分析及作图分别在 SPSS 23.0 和 Origin 2018软件中完成。
2 结果与分析
2. 1 4 种短命植物 C、N、P 含量及其计量比的动态变化特征
随着植物的生长发育,4种植物C、N、P含量及其化学计量比变化规律大致相似。由图 1可知,4 种植物 C 含量在整个生长期都保持相对稳定,且各生长期之间无显著差异(P>0.05)。4种植物 N、 P含量的变化趋势较为相似,均随生长期的延长呈现显著下降趋势,且差异显著(P<0.05)。化学计量比的动态变化显示(见图 2),4种植物的C∶N和 C∶P随着生长期的延长逐渐增大,且结实期和幼苗期、展叶期之间存在显著差异(P<0.05)。4种植物 N∶P呈现不规则变化,琉苞菊的N∶P在不同生长期存在显著差异(P<0.05),飘带果、假狼紫草和尖喙牻牛儿苗的N∶P仅在结实期变化显著(P<0.05)。
4 种植物 C、N、P 含量及其化学计量在整个生长期变化幅度不同(见表 1)。其中,C含量变化幅度最小,P 含量变化幅度最大。4 种植物 C 含量的变异系数均小于5%,为弱变异;而P含量的变异系数均大于 20%,为中等变异。假狼紫草的 N 含量变异系数(14.45%)小于 15%,为弱变异;其余 3 种植物 N 含量的变异系数则大于 15%,为中等变异。化学计量比中,C∶P 的变化幅度最大,其中尖喙牻牛儿苗(32.40%)、飘带果(30.31%),为中等变异。C∶N 比值的变化,尖喙牻牛儿苗最大,为 24.22%;假狼紫草最小,为 13.16%;飘带果和琉苞菊 为 15%~25%。 N∶P 比 值 的 变 化 ,除 飘 带 果(13.64%)和尖喙牻牛儿苗(13.88%)低于 15% 外,琉苞菊和假狼紫草的变异系数均大于15%,分别为 15.75%和18.19%。
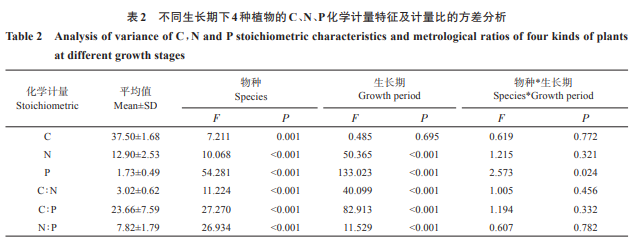
表2的双因素方差分析结果显示,同一生长期不同植物C含量存在显著差异(P<0.05),而同种植物在不同生长期C含量差异不显著(P>0.05),物种和生长期的交互作用对植物 C含量的影响也不显著(P>0.05)。植物的N、P、C∶N、C∶P、N∶P受物种、生长期影响均十分显著(P<0.05),但在两者的相互作用下仅P表现出显著差异(P<0.05)。
2. 2 荒漠土壤不同土层中 C、N、P含量及其化学计量比动态变化特征
从图 3 可以看出,荒漠土壤不同土层中 SOC、 TN、TP含量及其化学计量比随植物生长的变化不同。0~5 和 5~10 cm 土层的 SOC 含量随着植物生长期的延长逐渐增加,且自展叶期之后增幅迅速,而10~15 cm土层中SOC含量却呈现先增加后降低的趋势。3个土层中TN的含量随着植物生长期的延长先降低后增加,而 TP 含量却是先增加后降低,呈抛物线状变化趋势。TN和TP含量在植物结实期与幼苗期之间差异不显著(P>0.05)。5~10和 10~15 cm 土层的 SOC∶TN 在整个植物生长期内变化较大,表现为先上升后下降,且差异显著(P< 0.05),而0~5 cm土层土壤SOC:TN在整个生长期内基本持平,无明显变化(P>0.05)。SOC∶TP、TN∶TP 在3个土层中表现为较为相似的变化趋势,均在结实期达到最大值,且SOC∶TP在植物结实期与幼苗期差异显著(P<0.05)。
据表3可知,不同土层的土壤养分含量及变异系数有所不同。0~5 cm 土层土壤的 SOC、TN、TP 含量最高,分别达到 0.22、0.16和0.36 mg·g-1,且随土层深度加深含量逐渐减少。0~15 cm 土层土壤 SOC、TN、TP 的平均含量分别为 0.16、0.11 和 0.35 mg·g-1;化学计量比SOC∶TN、SOC∶TP、TN∶TP分别为1.63、0.45、0.31。土壤TP的变异性最弱,变异系数仅为 5.71%,土壤 SOC、SOC∶TP 的变异性较强,变异系数分别 31.25%、28.89%,为中等变异,土壤 TN、SOC∶TN、TN∶TP的变异系数均大于35%,表现为强变异。
表 4的双因素方差分析结果显示,不同土层、不同生长期土壤的SOC、TN、TP、SOC∶TN、SOC∶TP、 TN∶TP均存在显著差异(P<0.05)。土层和生长期的协同作用对土壤 SOC、SOC∶TN、SOC∶TP的影响显著(P<0.05),但 TN、TP、TN∶TP 在两者的协同作用下并无差异(P>0.05)。
2. 3 荒漠短命植物与土壤化学计量特征的相关关系
由表 5可知,4种短命植物与各层次土壤化学计量特征的相关性无一致规律。从单个植物来看,仅飘带果的 C与0~5 cm土层的 TP呈显著正相关(P<0.05),其它植物的化学计量特征与各土层 TP 均未表现出相关性。假狼紫草、尖喙牻牛儿苗的 N∶P 与 0~5 cm 土层的 TN∶TP 显著正相关,而飘带果的N∶P与10~15 cm土层的TN∶TP呈极显著正相关(P<0.01)。琉苞菊、假狼紫草、尖喙牻牛儿苗 的 C 与 0~5 cm 土 层 的 TN 显 著 负 相 关(P< 0.05),飘带果的P与0~5 cm土层的TN显著负相关(P<0.05)。基于不同土层分析,在 0~5 cm 土层,土壤 SOC、SOC∶TP与琉苞菊、假狼紫草、尖喙牻牛儿苗的C表现为显著负相关(P<0.05),与N表现 为显著正相关(P<0.05)。土壤 SOC 与四种植物的 P 呈极显著负相关(P<0.01),SOC、SOC∶TP 与四种植物的 C∶P、N∶P 呈显著正相关(P<0.05)。在 5~10 cm 土层,土壤 SOC、SOC∶TP 与飘带果、琉苞菊、假狼紫草的 N、P 表现为显著负相关(P< 0.05),而与 C∶N、C∶P 表现为显著正相关(P< 0.05)。在 10~15 cm土层,仅飘带果的 N∶P与该土层的 TN、TN∶TP 呈极显著正相关 (P<0.01),与 SOC∶TN呈显著负相关(P<0.05)。各土层土壤与植物化学计量总体相关性强度表现为 0~5 cm>5~ 10 cm>10~15 cm,且显著正相关多于显著负相关,说明植物在生长发育过程中,表层土壤对植物的促进及营养供给较为明显。
3 讨论
3. 1 4种短命植物化学计量特征的动态变化
C、N、P 等元素是植物进行光合作用、构成遗传物质的重要元素,为植物生长发育提供了良好的营养保障[19]。相关研究表明,植物种类的差异是造成其化学计量特征差异的关键因素之一[20]。本研究也发现,4 种植物的生理代谢受到生长期、植物种类因素的影响,体内化学计量特征存在较大的差异。而 4种短命植物不同生长期化学计量特征变化规律大致相似,这体现了同一生活型植物在资源分配利用方面的相对一致性。整个生长期内,C 元素的积累量最高,是由于 C 元素是构成植物结构的基本物质,总体需求量较大,同时 4种植物叶表面积大且具绒毛、芒刺等附属物质,光合作用合成有机物能力强[5],故C元素在植物体内含量高。N、P 元素从幼苗期到结实期逐渐减少,原因可能是在幼苗期,植物体生物量小且处于快速生长发育阶段,需要吸收较多 N、P 元素合成大量的核酸蛋白质进行细胞分裂,因而体内 N、P 元素浓度较高,随着植物的不断生长,生物量逐渐增大从而使 N、P 元素含量相对降低,这与牛得草等[21]的研究结果相似。
植物的化学计量比能够清晰地表明植物营养缺失情况。C∶P和 C∶N代表植物同化碳能力的强弱,能够体现其养分利用效率的高低,具有至关重要的生态学意义[22]。本研究中,飘带果的 C∶N 最高,假狼紫草的C∶P最高,显示出这两种植物在营养贫瘠的古尔班通古特沙漠具有较强的固碳能力和养分利用效率。N∶P的大小常被用作植物生长受限元素的判断,N∶P<14.0为缺N,N∶P>16.0为缺P [23]。本研究中 4种植物不同生长期的 N∶P(5.26~ 12.88)均显著低于 14,表明该区域短命植物生长的限制元素为氮素,与刘建国等[17]的研究结果相似。4种植物N∶P均在生长末期突然增大,可能与该地区五月份时干旱无雨有关,干旱会导致植物体加强相应的保护系统,促使叶片内可溶性蛋白提高来缓解干旱胁迫,而植物体内 N 元素的主要存在形式是可溶性蛋白,故 N∶P 相对得到了提高[24]。通过对 4种植物 C、N、P含量及计量比在整个生 长期的综合变异分析,变异系数由大到小依次为:C∶P>P>C∶N>N>N∶P>C,C的变异系数最小, N、P 较大,表明该荒漠区植物 C 含量相对稳定,而 N、P 含量具有较大的季节变异。本研究中 4 种植物N、P、N:P的变化范围均在李玉霖关于北方荒漠区 214 种植物叶片 N、P、N∶P 的研究报道之内[25],但 N、P、N∶P的变异系数排序与李玉霖的报道(P> N∶P>N)稍有不同,可能是本研究植物样品采集时间与其不同,且样品采集尺度较小,所以,今后的研究有必要跨越植物整个生长季进行大时间尺度的样品采集。
3. 2 不同土层荒漠土壤化学计量特征分析
SOC、TN、TP含量及其化学计量比是衡量土壤有机质组成和质量的关键性指标[26]。本研究发现,土壤表层(0~5 cm)SOC、TN、TP平均含量最高,且随土壤深度增加呈下降趋势,与陶冶等[27]研究结果相似。0~15 cm土壤SOC、TN、TP平均含量分别为0.16、0.11和0.35 mg·g-1,土壤SOC、TN、TP整体含量明显低于全国均值水平(11.21、1.06、0.65 mg·g-1)[28]。化学计量比 SOC∶TN、SOC∶TP、TN∶TP(1.63、0.45、 0.31)虽能很好地诠释土壤养分状况,但也远远低于全国均值(12.01、25.77、2.15)[28],这可能是受气候、海拔、地貌、植被、降水等因素的影响,造成该荒漠区土壤养分元素极为匮乏。从变异系数来看,TP 变异系数最小(5.71%),而 SOC、TN 则较大(16.67%、36.36%),是由于 SOC、TN 主要受土壤基质与枯落物的分解以及植物吸收利用的影响,存在较大的空间变异性,而 TP主要受土壤基质的影响[29],产生的差异相对较小,这也表明 3种土壤化学计量比主要受控于SOC和TN含量。一般认为,降雨会导致土壤TP的淋溶作用增强,不利于TP的累积[30~31],而研究区土壤TP含量较低,可能是由于沙漠区总体上降雨量少,矿物质的风化以物理风化为主,造成P含量相对较低。
SOC∶TN可以影响土壤中碳氮元素的循环,是衡量土壤质量的关键性指标。SOC∶TN 的大小与有机质分解速度呈反比,即SOC∶TN较低的土壤具有较快的矿化作用[32]。相比于全国平均水平的 SOC∶TN(14.3)[28],该研究区的SOC∶TN(1.63)低于全国平均水平,表明研究区土壤有机质分解和矿化速率相对较高。在不同土层和时间梯度下,土壤 SOC∶TN 始终维持相对稳定的水平,表明 SOC、 TN 元素之间存在显著的相关关系,SOC、TN 元素对土壤微环境变化的响应几乎是同步的,同时也验证了生态系统中土壤 SOC∶TN 保持相对稳定的理论[33]。土壤 SOC∶TP 的高低显著会影响植物的生长发育,是评价土壤 TP 矿化能力高低的标志,也是衡量微生物矿化土壤有机质释放 TP的指标。较低的 SOC∶TP 有利于微生物分解有机质释放养分,反之,土壤 SOC∶TP 过高则会导致微生物在分解有机质的过程中受到 TP 的限制从而与植物竞争土壤中的 TP,进而不利于植物吸收养分[34]。研究区土壤 SOC∶TP(0.45)远远低于全国平均值(9.3)[28],表明该研究区土壤 TP 具有较高的有效性,土壤中的微生物分解过程中不再受到 TP的限制。TN∶TP常被用于确定土壤养分限制元素的阈值[31]。本研究发现,该研究区具有较低的 N 含量以及 TN∶TP 比,这表明本研究区土壤 N 元素相当贫瘠,也进一步佐证了该区域短命植物生长的限制元素为N素。——论文作者:王明明 庄伟伟*
澹版槑:鈶犳枃鐚潵鑷煡缃戙€佺淮鏅€佷竾鏂圭瓑妫€绱㈡暟鎹簱锛岃鏄庢湰鏂囩尞宸茬粡鍙戣〃瑙佸垔锛屾伃鍠滀綔鑰�.鈶″鏋滄偍鏄綔鑰呬笖涓嶆兂鏈钩鍙板睍绀烘枃鐚俊鎭�,鍙仈绯�瀛︽湳椤鹃棶浜堜互鍒犻櫎.

SCISSCIAHCI

