ЗЂВМЪБМфЃК2022-02-15ЫљЪєЗжРрЃКХЉвЕТлЮФфЏРРЃК1ДЮ
еЊ вЊЃК еЊ вЊЃКЕзЦмгаПзГцдкЙХЛЗОГбаОПжаЕФгІгУЯШЪЧгУзїЙХЫЎЩюЛђЫЎЭХЕФБъжОЮяЁЃЫцзХаТММЪѕЕФгІгУ ЪЖБ№ГіСЫ2жжВЛЭЌЕФЕзЦмгаПзГцЮЂЩњОГЃКЭтЩњжжКЭФкЩњжж;ШЯЪЖЕНЩѕжСЩюКЃЕзЦмгаПзГцвВФмЖдБэВуИЁгЮЩњЮяВЊЗЂЕФМОНкадЖЬднЪТМўзїГіЯьгІвђЮЊГСНЕЕНКЃЕзЕФгаЛњЮяжЪЙЉбјзХЕзЦмгаПзГцЁЃФП
ЁЁЁЁеЊ вЊЃКЕзЦмгаПзГцдкЙХЛЗОГбаОПжаЕФгІгУЯШЪЧгУзїЙХЫЎЩюЛђЫЎЭХЕФБъжОЮяЁЃЫцзХаТММЪѕЕФгІгУ‚ ЪЖБ№ГіСЫ2жжВЛЭЌЕФЕзЦмгаПзГцЮЂЩњОГЃКЭтЩњжжКЭФкЩњжж;ШЯЪЖЕНЩѕжСЩюКЃЕзЦмгаПзГц‚вВФмЖдБэВуИЁгЮЩњЮяВЊЗЂЕФМОНкадЖЬднЪТМўзїГіЯьгІ‚вђЮЊГСНЕЕНКЃЕзЕФгаЛњЮяжЪЙЉбјзХЕзЦмгаПзГцЁЃФПЧА‚ЕзЦмгаПзГцБЛЙуЗКгІгУгкЙРЫуКЃбѓБэВуЩњВњСІКЭЕзВуЫЎЭХЕФКЌбѕСПЁЃЛиЙЫСЫЩюКЃЕзЦмгаПзГцЩњЬЌбаОПЕФРњЪЗКЭЦфдкЙХКЃбѓбЇжаЕФгІгУ‚ВЂЧПЕїбаОПЁЂгІгУжаЕФаТЗНЗЈЁЂаТММЪѕЁЃжаЙњвбОМгШыСЫжюШч IODP ЕШЩюКЃбаОПМЦЛЎ‚гаБивЊЯђЮвЙњбЇЪѕНчЬсЙЉЕзЦмгаПзГцбаОПЕФаТЗНЯђ‚вдзЪВЮПМЁЃ
ЁЁЁЁЙи Мќ ДЪЃКЕзЦмгаПзГц;ЩюКЃбаОП;гаЛњЬМЭЈСП;КЌбѕСП;злКЯДѓбѓзъЬН
ЁЁЁЁЕзЦмгаПзГцЪЧЮЂЬхЙХЩњЮябЇЕФЦ№ЕуЁЃЕквЛДЮЪРНчДѓеНЧАКѓ‚УРЙњПЦбЇМвЪзЯШНЋетРрЮЂЬхЛЏЪЏзїЮЊЕиВуФъДњЕФБъжОгУгкЪЏгЭПБЬН‚ДѓЛёГЩЙІ‚ДгЖјНЈСЂСЫЮЂЬхЙХЩњЮябЇетУХаТбЇПЦЁЃжСгкИЁгЮгаПзГцЕФЕиВувтвх‚вЊЕН20ЪРМЭ50ФъДњВХБЛЗЂЯж‚ИЦжЪГЌЮЂЛЏЪЏЕФгІгУИќЭэЁЃдкЯжДњЕФЕижЪПЦбЇжа‚ЮЂЬхЛЏЪЏЪзЯШЪЧЙХКЃбѓбЇЁЂЙХЛЗОГбаОПЕФЪжЖЮ‚ИЁгЮгаПзГцКЭИЦжЪГЌЮЂЛЏЪЏЕФгІгУБШЕзЦмгаПзГцИќЮЊЙуЗКЁЃЪЧВЛЪЧЕзЦмгаПзГцвбО“ЭЫОгЖўЯп”‚дкЩюКЃбаОПжаВЛФЧУДживЊСЫФи?
ЁЁЁЁБОЮФДгРњЪЗЛиЙЫШыЪж‚ЗжЮіЕзЦмгаПзГцбаОПМАЦфгІгУЕФБфЛЏРњГЬ‚жИГіЦфдкЩюКЃбаОПжаВЛПЩЬцДњЕФживЊадЁЃВЛЪЧЕзЦмгаПзГцЕФбаОПМлжЕЗЂЩњСЫБфЛЏ‚ЖјЪЧЫќЫљНтЕФПЦбЇЮЪЬтКЭбаОПЕФЗНЗЈвбОзЊвЦ‚ жЕЕУЮвЙњЕибЇНчЁЂгШЦфЪЧЙХКЃбѓбЇНчзЂвтЁЃ
ЁЁЁЁ1 ЕзЦмгаПзГцзїЮЊ“БъжОжж”
ЁЁЁЁЕзЦмгаПзГцдкдчЦкЕФЩюКЃбаОПжа‚КЭЦфЫќЙХЩњЮяЛЏЪЏгІгУгкЕиВубЇЛђЙХЩњЬЌбЇЪБзїЮЊ“БъжО”ЮявЛбљ‚ГЃГЃЪЧзїЮЊЩюЖШЛђепЫЎЭХЕФБъжОЁЃ
ЁЁЁЁдчдк20ЪРМЭСљЦпЪЎФъДњвдЧА‚вбОЗЂЯжСЫЩюКЃЕзЦмгаПзГцзщКЯгыЩюЖШЯрЙи‚ЫфШЛФЧЪБЖдЦфЩњЬЌСЫНтВЛЖр‚ЕЋЦеБщАбЕзЦмгаПзГцгУзїЙХЩюЖШБъжО[1]‚ ШЯЮЊ“ЕзЦмгаПзГцШКЪЧдкАЖЭтАДЫЎЩюГЪДјзДЗжВМ” [2]ЁЃЮвЙњдкбаОПКЃЧјЕзЦмгаПзГцЩњЬЌЗжВМЕФЙ§ГЬжа‚вВЪзЯШзЂвтЕНгыЩюЖШЕФЙиЯЕЁЃКѓРДЕФбаОПБэУї‚ЕзЦмгаПзГцЕФЩюЖШЗжВМ‚КмДѓГЬЖШЩЯЪЧЫЎЭХгыЫЎЩюЕФЙиЯЕ‚дкВЛЭЌЩюЖШДІЗжВМзХВЛЭЌЕФЫЎЭХ‚БЛВЛЭЌЫЎЭХеМОнЕФКЃЕзЩњЛюзХВЛЭЌЪєжжзщКЯЕФЕзЦмгаПзГцЁЃвђДЫ‚дкбаОПЙХЩюЖШЪБ‚вЊЧѓЖдЙХЫЎЬхзізлКЯадНтЪЭ[3]‚ВЛФмгУвЛИіКЃЧјЕФЩюЖШзЪСЯжБНгШЅНтЪЭСэвЛКЃЧјЕФЙХЩюЖШ[4]ЁЃ
ЁЁЁЁгыДЫЭЌЪБ‚ЕзЦмгаПзГцЕФ“БъжОжж”вВБЛЙуЗКгУгкжИЪОЫЎЭХ‚БШШчдкДѓЮїбѓЩюЫЎЧј‚Epistominella umbonifera жИЪОФЯМЋЕзВуЫЎ‚Epistominella exigua КЭ Cibicidoides w uellerstorf i жИ ЪО ББ Дѓ Юї бѓ Щю ВуЫЎ[5]ЁЃгыЫЎЩюЯрБШ‚ЫЎЭХДњБэЕФЪЧЖржжЛЗОГвђЫиЕФзлКЯ‚ЖјВЛжЛЪЧЕЅвЛЕФЛЗОГвђЫи[6]‚НЋЕзЦмгаПзГцгІгУгкЫЎЭХбаОПЯдШЛЪЧвЛжжНјВНЁЃЕЋЮоТлЫЎЩюЛђепЫЎЭХ‚ЕзЦмгаПзГцжБЕН80ФъДњГѕЛЙжЛЪЧзїЮЊ“БъжОжж”дкЪЙгУЁЃОЁЙмгУЛЂКьШОЩЋЪЖБ№“Лю”гаПзГц(ЦфЪЕЪЧМьВщгаЮодЩњжЪ)РДбаОПЩюКЃЕзЦмгаПзГцЩњЬЌЕФММЪѕдчдк50ФъДњОЭвбОЗЂУї‚ЕЋгЩгкВЩбљКЭЙлВтММЪѕЕФЯожЦ‚етжжЩњЬЌбаОПдкЕБЪБЛЙжЛФмЪЧОВЬЌЕФЁЂЕЭЗжБцТЪЕФ‚гыНёЬьвдЖЈСПЗжЮіЮЊЛљДЁЁЂвдзЗЫїЛњРэЮЊФПБъЕФЙХКЃбѓбЇвЊЧѓЯрБШ‚БуЯдЕУВЛЯрЪЪгІЁЃ
ЁЁЁЁ2 ЩюКЃЕзЦмгаПзГцЮЂЩњОГ
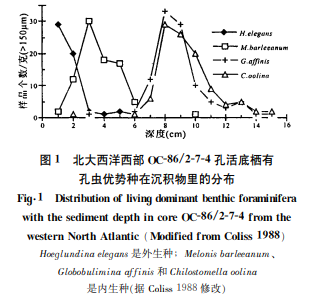
ЁЁЁЁ80ФъДњЦ№‚КЃЩЯММЪѕЕФЗЂеЙЮЊЕзЦмгаПзГцЩњЬЌбаОППЊБйСЫаТНзЖЮЁЃУРЙњ Corliss [7]АбЯфЪНШЁбљЦїШЁЩЯЕФ OC-86/2-7-4ПзЩюКЃГСЛ§жљзДбљЧаГЩ1cm КёЕФБЁВу‚дкДЌЩЯЗжБ№гУЛЂКьШОЩЋ‚НсЙћЗЂЯжЕзЦмгаПзГцВЛНіЗжВМдкКЃЕзГСЛ§ВуЕФБэУц‚ЛЙЩњЛюдкГСЛ§ЮяжЎжа‚ДЙЯђЩЯГЪВузДЗжВМ(ЭМ1)‚жЄЪЕСЫЫћвдЧАИљОнОнЕзЦмгаПзГцПЧЬхЕФЬМЭЌЮЛЫиВювьЭЦВтФГаЉЕзЦмгаПзГцПЩФмЩњЛюдкГСЛ§ЮяРяЕФЩшЯы‚вђЖјЬсГіСЫЕзЦмгаПзГцЮЂЩњОГ(microhabitat)ЕФИХФю‚НЋЕзЦмгаПзГцЧјЗжЮЊЭт(Бэ)ЩњжжКЭФкЩњжжСНДѓРр(ЪЕМЪЩЯЛЙДцдквЛРрНщгкЭтЩњжжКЭФкЩњжжжЎМфЕФЛњЛсжж)ЁЃ
ЁЁЁЁгаЙиЮЂЩњОГЕФбаОПЪЧЕзЦмгаПзГцЩњЬЌбЇЕФвЛИіживЊНјеЙЁЃгывЛАуЩњЮяЕФЩњОГИХФюВЛЭЌ‚ЕзЦмгаПзГц“ЮЂЩњОГ”ЩцМАЕФЪЧКЃЫЎгыГСЛ§ЮяНчУцЩЯКЭНчУцвдЯТГСЛ§ЮяРяЕФДЙЯђЗжВМ‚ЧПЕїгаПзГцЛюЖЏЕФЯрЖдЮЛжУЁЃдк Coliss [7]ЗЂЯжЕзЦмгаПзГцдкКЃЕзГСЛ§ЮяРяГЪВузДЗжВМЕФЭЌЪБ‚ЫћЛЙЗЂЯжСЫПЧЬхаЮЬЌгыЮЂЩњОГЕФЙиЯЕЃКОпгаа§ОэаЭПЧЬхЕФЕзЦмгаПзГцжа‚БэЩњжжОпгаТна§ПЧКЭЫЋЭЙЛђЦН-ЭЙЕФПЧаЭ‚БугкдкГСЛ§ЮяБэУцБЃГжЮШЖЈ;ФкЩњжжЭљЭљОпгаЦНа§ПЧКЭЛыдВЕФПЧдЕ‚БугкдкГСЛ§ВуФкВПвЦЖЏЁЃгЩгкГСЛ§ВуБэВуКЭФкВПКЌбѕСПВЛЭЌ‚БэЩњжжПЧЬхЕФЯИПзПЩвджЛдкБГУцЗЂЩњ‚ЖјФкЩњжжЕФЯИПздђдкПЧУцОљдШЗжВМ‚вдЪЪгІдЩњжЪгыЭтНчЦјЬхНЛЛЛЕФашвЊЁЃ
ЁЁЁЁВЛНіШчДЫ‚Corliss [8]дкХВЭўКЃЕФбаОПНсЙћБэУї‚ ЕзЦмгаПзГцЕФаЮЬЌЬиеїгыЩюЖШЗжВМОпгаЯрЙиад(ЭМ 2)‚ЖјетжжЯрЙиадЪЕМЪЩЯЗДгГСЫЮЂЩњОГгыЩюЖШЕФЙиЯЕЁЃПЧЬхаЮЬЌЪЧзЖаЮЁЂдВжљаЭЁЂБтЦНзЖаЭЁЂЛыдВЦНа§аЭЁЂдВЧђаЭЛђБтЦНЭждВаЭЕФФкЩњжж‚ЧїЯђгкЗжВМдк 500ЁЋ1000m жЎМфЯрЖдЧГЕФКЃЕз;ПЧЬхаЮЬЌЪЧЦН-ЭЙТна§аЭЁЂЫЋЭЙТна§аЭЁЂЫкзДТна§аЭКЭЛыдВТна§аЭЕФЭтЩњжж‚ЧїЯђгкЗжВМдк1500m вдЯТЕФКЃЕз;1000 ЁЋ1500m дђЪЧЖўепжЎМфЕФЙ§ЖЩ‚200ЁЋ500m ДІПЩФмвђЖўепЪмЩњЮяШХЖЏЖјКЌСПЛљБОЯрЕБЁЃФЋЮїИчЭхЕФбаОПНсЙћгыХВЭўКЃЕФбаОПНсЙћЛљБОвЛжТЃК1300m вдЩЯФкЩњжжеМжїЕМЕиЮЛ‚2000m вдЯТжївЊЪЧЭтЩњжж‚1300ЁЋ2000 m ЪЧФкЩњжжКЭЭтЩњжжжЎМфЕФЙ§ЖЩ[9]ЁЃЮЂЩњОГгыЩюЖШЕФЙиЯЕ‚гжШЁОігкгаЛњЬМЕФЭЈСПЃКгаЛњЬМЭЈСПИпЕФКЃЕз‚вдФкЩњжжЮЊжї‚ЖдгІЕФКЃЫЎЩюЖШНЯЧГ;гаЛњЬМЭЈСПЕЭЕФКЃЕз‚ЫЎЩюНЯДѓ‚вдЭтЩњжжЮЊжї‚вВОЭЪЧЫЕ‚гаЛњЬМЭЈСППижЦзХЕзЦмгаПзГцЭтЩњжжКЭФкЩњжжЕФБШР§ЁЃЦЉШч‚ХВЭўКЃгаЛњЬМЭЈСПКЭЮЂЩњОГЕФБфЛЏвд1000m ЮЊНч(ЭМ2)ЁЃ
ЁЁЁЁЮЂЩњОГЕФСэвЛИіЗНУцЪЧКЃЕзЕФбѕЛЏЛЙдзДПіЁЃБэЩњжжгыФкЩњжжЩцМАСНРрВЛЭЌЕФШмНтбѕЃККЃЕзЫЎЬхЕФШмНтбѕКЭКЃЕзГСЛ§ЮяПзЯЖЫЎРяЕФШмНтбѕЁЃКЃЕзЫЎЬхЕФШмНтбѕХЈЖШЪЧЩюВПЫЎЭХЕФвЛИіживЊаджЪ‚ЖдгкИќКУЕиРэНтКЃбѓЛЗСїРњЪЗЁЂЦјКђБфЛЏЕШОпгаживЊвтвхЁЃПзЯЖЫЎРяЕФШмНтбѕХЈЖШЖдгаПзГцЕФПЧБкЬиеїМАЦфЩњЛюЕФЮЂЩњОГгаживЊгАЯь‚ФкЩњжжПЧЬхжЛгаБШЭтЩњжжБкПзУмЖШДѓЕУЖр‚ВХФмдіЧПгыЭтНчЕФЦјЬхНЛЛЛ[7]ЁЃгаЙиШмНтбѕЖдгаПзГцгАЯьЕФбаОП‚вд Kaiho [10]ЕФЙЄзїзюОпДњБэад‚ЫћНЈСЂСЫИЦжЪЕзЦмгаПзГцбѕХЈЖШжИЪ§‚ШЯЮЊШмНтбѕХЈЖШЯожЦзХЕзЦмгаПзГцЕФЮЂЩњОГЃКдкбѕЛЏЛЗОГ(oxic‚>1.5mL/L O2)Ря‚ЕзЦмгаПзГцвдЭтЩњжжеМгХЪЦ‚ОпгаПЧЬхКёЁЂПЧОЖДѓ(≥350 μm)ЕФаЮЬЌЬиеї;дкШѕбѕЛЏЛЗОГ(dysoxic‚0.1ЁЋ0.3 mL/L O2)Ря‚ЕзЦмгаПзГцзщГЩжївЊЪЧФкЩњжж‚ОпгаПЧЬхБЁЁЂПЧОЖаЁ(Тна§ПЧ<200μm‚ЩьГЄБтЦНПЧ< 250μm)ЕФЬиЕу;ЮЈЖРдкДЮбѕЛЏЛЗОГ(suboxic‚0.3ЁЋ 1.5 mL/L O2)Ря‚ЭтЩњжжКЭФкЩњжжПЩвд“КЭЦНЙВДІ”‚ЕЋвВвЊвдМѕаЁИіЬхДѓаЁ(<350μm)ЮЊДњМлЁЃбѕЛЏЛЗОГРяЕФЭтЩњжжКЭШѕбѕЛЏЛЗОГРяЕФФкЩњжжЩњЛюдкбѕКЌСПИп(>2mL/L O2)ЕФЕзВуЫЎЯТ;ЖјДЮбѕЛЏЛЗОГЕФгаПзГцвЛАудкЕЭбѕЕзВуЫЎжаГЃМћЁЂЕЋдкИпбѕЕзВуЫЎЛЗОГРяКЭНгНќШБбѕЛЗОГРявВПЩвдЩњДцЁЃKaiho РћгУЩюКЃЕзЦмгаПзГцРДбаОПШмНтбѕКЌСПЪЧЩюВПЫЎЭХаджЪбаОПжаЕФвЛИіЕфЗЖЁЃ
ЁЁЁЁЮЂЩњОГЕФбаОП‚ЕМжТСЫПижЦвђЫижЎељ[11]ЃКВПЗжбЇепШЯЮЊКЌбѕСПЪЧПижЦвђЫи‚ЖјСэвЛВПЗжбЇепдђШЯЮЊгЊбјЮяЙЉгІЪЧзюживЊЕФПижЦвђЫиЁЃНќФъРДЖдКЃбѓБэВуЩњВњСІгыЕзЦмгаПзГцЙиЯЕЕФбаОП‚ЮЊЛиД№етИіЮЪЬтЬсЙЉСЫживЊвРОнЁЃ
ЁЁЁЁ3 гЊбјЮяжЪЙЉгІгыЕзЦмгаПзГц
ЁЁЁЁЩюКЃЕзЦмгаПзГцЕФЪГЮяРДдДгыЦфЩњДцЛЗОГУмЧаЯрЙиЁЃЩюКЃЕзЦмгаПзГцДѓжТЩњЛюдкСНРрЛЗОГРяЃКШЋФъЕЭЮТЁЂИпЫЎбЙЁЂЭъШЋКкАЕЕФвЛАуЩюКЃЛЗОГКЭгаЩюКЃШШвКЁЂРфЫЎЩјЭИЕШЕФЬиЪтЛЗОГ[12]ЁЃдквЛАуЩюКЃЛЗОГРя‚КЃбѓБэВуЩњВњСІЪЧЕзЦмгаПзГцЕФгЊбјЮяжЪЬсЙЉеп‚гЊбјЮяжЪЙЉгІЭљЭљЪЧМОНкадБфЛЏЕФ;ЖјдкЬиЪтЛЗОГРя‚гЊбјЮяжЪЙЉгІЪЧЗЧМОНкадЕФ‚ЪГЮяРДдДвВКмЖрбљ‚гыЧАепЯрБШ‚ШЫУЧЖдЦфШЯЪЖЛЙКмЗєЧГЁЃ
ЁЁЁЁ3.1 МОНкадЪГЮяЙЉгІЖдЕзЦмгаПзГцЕФгАЯь
ЁЁЁЁКЃбѓБэВуЩњВњСІЪЧЕзЦмгаПзГцгЊбјЮяжЪЕФжївЊЬсЙЉеп‚ДгКЃбѓБэУцГСНЕжСКЃЕзЕФгЊбјЮяжЪЮЌГжзХвЛАуЩюКЃЛЗОГРяЕФЕзЦмгаПзГцжжШКЕФЩњДцКЭЗБбм‚ ГСЕНКЃЕзЕФгаЛњЬМдНЖр‚гаПзГцЗсЖШвВОЭдНИпЁЃдјОЭЦЫу‚УП1mg ЕФгаЛњЬМНЕЕНКЃЕз‚ОЭПЩвдгавЛИіПЧОЖ>150μm ЕФЕзЦмгаПзГцЩњГЩ(вђДЫЬсГігУЕзЦмгаПзГцЕФЖбЛ§ЫйТЪЙРЫуЙХЩњВњСІ[13])ЁЃгЩгкКЃУцдхРрВЊЗЂКѓГСНЕЕНКЃЕзЕФжВЮяЫщаМЯШГЪИЁНЌ(fluff)зДДцдк‚етРрИЁНЌОЭЮЊзЗзйЕзЦмЩњЮяЖдБэВуЩњВњСІЪТМўЕФЯьгІЬсЙЉСЫВФСЯЁЃЙћШЛ‚80ФъДњжаЦк‚гЂЙњ Gooday дкББДѓЮїбѓ4000m зѓгвЫЎЩюДІВЩМЏЕНКЃЕзИЁНЌ‚ДгжаЗЂЯжСЫДѓСПЕФЕзЦмгаПзГцЛюИіЬх‚Цфжавд Epistominella exigua ЕШ3ИіжжеМОнгХЪЦ[14]ЁЃетЯюЗЂЯжИФБфСЫЩюКЃЕзЦмгаПзГцЖдКЃбѓБэВуЛЗОГЗДгГВЛСщУєЕФЦЋМћ‚жЄУїЛюгаПзГцШКдкКЃбѓЕзВПвВФмЯьгІМОНкадБфЛЏКЭБэВуКЃЫЎЕФЪТМў[15]ЁЃ
ЁЁЁЁИќМгЯЕЭГЕФбаОПРДздШеБОЁЃKitazato ЕШ[12]дкЖЋОЉвдФЯЕФЯрФЃЭхЧЇгрУзЕФЩюЫЎДІ‚НјааСЫ4Фъ(1994-1998)ЕФЖЈЕуЙлВт‚ВЩгУвЃВтвЃИа(КЃбѓЩњВњСІ)ЁЂГСЛ§ВЖМЏЦї(ГСНЕПХСЃ)ЁЂКЃЕзТМЯё(ЮэзДВу[nepheloid layer]ЙлВт)КЭЖрЙмШЁбљ(ГСЛ§ЮяМАБэУцЕФИЁНЌ)ЕШЖржжЪжЖЮ‚ЗЂЯжКЃУцдхРрВЊЗЂ(ЭМ3A)Кѓ‚ВЛНіГСЛ§ЭЈСПдіДѓ(ЭМ3B)‚ЖјЧвКЃЕзЕФЮэзДВуБфХЈ(ЭМ 3C)‚ИЁНЌВудіКё(ЭМ3D)‚ГСЛ§ЮябѕЛЏВуМѕБЁ(ЭМ3E)‚ ЛюЕзЦмгаПзГцвВЯргІдіЖр(ЭМ3F)‚ЩњЖЏЕиеЙЯжСЫБэВуЩњВњСІЧ§ЪЙЕзЦмгаПзГцЗБЪЂЕФЙ§ГЬЁЃ
ЁЁЁЁ90ФъДњвдРД‚ИЁгЮЩњЮяВЊЗЂЪТМўЖдЕзЦмгаПзГцЕФгАЯь‚вбОв§Ц№бЇЪѕНчЕФУмЧаЙизЂ‚ЪЙЕУгаЙиБэВуЩњВњСІЛђепгаЛњЬМЭЈСПгыЕзЦмгаПзГцЪєжжзщКЯЕФЙиЯЕЧїгкУїРЪЁЃдкЩњЮяВЊЗЂЦкМф‚ГСНЕжСКЃЕзЕФгаЛњЬМзмСПеМФъгаЛњЮязмСПЕФДѓВПЗж[16]‚ВЂЧвЫќУЧЖрВЛЮШЖЈ‚НЕНтПьЁЃЕзЦмгаПзГцЪЪгІетжжЛЗОГ‚ЬиЛЏГіСЫЖЬЦкбИЫйЗБжГЩњГЄКЭГЄЦкШЬЪмМЂЖіЕФФмСІ‚ДгЖјВњЩњГіЫцгаЛњЬМЭЈСПБфЛЏЕФЪєжжзщКЯЃКБэЩњжжКЭНКНсжЪЧГФкЩњжжЪЧЕЭгаЛњЬМЭЈСПКЃЕзЕФЬиеїзщКЯ;ЖјдкИпгаЛњЬМЭЈСПЕФКЃЕз‚гаЛњЛсжж(вВНаднЪБадБэЩњжж)КЭФкЩњжжзщКЯ(ЭМ4)ЁЃВЛНіШчДЫ‚ВЛЭЌКЃЧјЕФБэВуЩњВњСІМОНкадБфЛЏЭљЭљКмВЛвЛбљ‚ЦфЧПШѕКЭгаЛњЮяжЪЯђКЃЕзМфаЊЪНГСЛ§ЕФЦЕТЪвВЛсгАЯьЕзЦмгаПзГцЪєжжзщКЯ[17]ЁЃ
ЁЁЁЁгаЙигаЛњЬМЭЈСПгыЕзЦмгаПзГцзщКЯЙиЯЕЕФаТШЯЪЖ‚ЪЧЖд Coliss ЕШ[8]ЗЂЯжЕФдкВЛЭЌЫЎЩюДІгаЛњЬМЭЈСПКЭЮЂЩњОГЙиЯЕЕФНјвЛВНЗЂеЙ‚вВЮЊгаПзГцзщКЯЕФЩюЖШЗжВМЬсЙЉСЫНтЪЭЃКВЛЭЌЩюЖШКЃЧјЕФЩњВњСІКЭгаЛњЬМЭЈСПВЛЭЌ‚ЕМжТСЫЕзЦмгаПзГцЪєжжЗжВМЕФВЛЭЌЁЃвдЕижаКЃЮЊР§‚БэВуЩњВњСІгЩЮїЯђЖЋЕнМѕ‚вђДЫЭЌвЛЩюЖШЕФЕзЦмгаПзГцзщКЯвВГЪЖЋЮїЯђБфЛЏ;ЖјЭЌвЛжжЕФЩюЖШЗжВМНчЯовВЫцзХгаЛњЬМЭЈСПЕФВЛЭЌЖјБфЛЏ[20]‚ЦЉШчЭтЩњжж Gyroidina orbicularis ЩњЛюдкНЯЩюКЃЕз‚еМШЋШКТф10%вдЩЯЕФИпжЕЧјЩЯЯо‚дкЮїЕижаКЃЮЊ1500m‚ЯђЖЋдђЩЯЩ§ЕН800ЁЋ600mЁЃ
ЁЁЁЁ3.2 ЗЧМОНкадЪГЮяЙЉгІЖдЕзЦмгаПзГцЕФгАЯь
ЁЁЁЁдкЩюШыбаОПБэВуЩњВњСІМОНкадБфЛЏЖдЩюКЃЕзЦмгаПзГцгАЯьЕФЭЌЪБ‚ЗЧМОНкадЪГЮяЪфШыЖдЩюКЃЕзЦмгаПзГцЕФгАЯьвВв§Ц№СЫШЫУЧЕФзЂвтЁЃгЩгкЗЧМОНкадЪГЮяЪфШыЕФжжРрНЯЖрЧвНЯИДдг‚ФПЧАЖдЦфКЭЕзЦмгаПзГцЕФЙиЯЕЛЙСЫНтВЛЖр‚ЯёОЈЪЌЁЂЮоМЙзЕЖЏЮяЁЂФОВФЕШгЊбјЮяжЪЯђКЃЕзЕФЪфШыУЛгаЙцТЩ‚вВВЛПЩдЄВт‚дкДѓЖрЪ§ЕиЗНОпгаОжВПад‚ЫќУЧЖдЕзЦмгаПзГцЕФгАЯьгыздЩэДѓаЁКЭаджЪгаЙи;ЖјдкЕЭЧПЖШзЧСїЦЕЗБЗЂЩњЕФЕиЗН‚ЕзЦмгаПзГцФмЙЛвРППгаЛњЮяжЪГжајЩњДц;дкгаЩюКЃШШвКЩјЭИЕФКЃЕз‚гаПзГцПЩФмЪЧЕБЕиЖЏЮяШКЕФживЊзщЗж‚ЦфУмЖШЁЂЖрбљадЁЂзщГЩЖрВЛЭЌгкЗЧШШвКЧјЖЏЮяШК[16]ЁЃвЛАуЫЕРД‚ЪЪгІЗЧМОНкадгЊбјЙЉгІЕФгаПзГцЭљЭљЪЧФЧаЉФмЪЪгІЕЭбѕКЭ(Лђ)ИпЩњВњСІЛЗОГЕФЮяжж‚ЫќУЧЯьгІгаЛњЮяЪфШыЕФЗНЪНПЩФмвђгаЛњЮяБОЩэЕФаджЪВЛЭЌКЭЪфШыЕФЙцФЃКЭЪБМфГпЖШВЛЭЌЖјгаВюБ№[18]ЁЃ
ЁЁЁЁзмжЎ‚ЩюКЃбаОПжаЕФЕзЦмгаПзГцвбгЩЕБГѕЕФЙХЩюЖШКЭЫЎЭХЕФ“БъжОжж”‚БфГЩСЫФмЙЛНвЪОШмНтбѕКЌСПКЭБэВуЩњВњСІБфЛЏ(вВФмЗДгГгаЛњЮяЗЧМОНкадБфЛЏ)ЕФ“вЊдБ”‚ГЩЮЊвЛЩэМцЖржАЕФ“ЖрУцЪж”ЁЃетЙЬШЛЫЕУїЦфгІгУЗЖЮЇБфПэ‚ЕЋвВБъжОзХЖдЕзЦмгаПзГцбаОПГЬЖШЕФЩюШыЁЃЪЕМЪЩЯ‚КЃЫЎЩюЖШ(ЛђЫЎЭХ)ЁЂгаЛњЬМЭЈСПЁЂШмНтбѕКЌСПжЎМфДцдкФГжжЙиСЊадЁЃдкЗжВМзХВЛЭЌЫЎЭХЕФКЃЫЎЩюЖШДІ‚гаЛњЬМЭЈСПЭљЭљвВВЛвЛбљ‚ ЖјгаЛњЬМЭЈСПгыШмНтбѕКЌСПЭЈГЃГЪЯжИКЯрЙиадЁЃЫќУЧжЎМфЕФЙиЯЕЪЕМЪЩЯЪЧКЃЫЎЩюЖШгыЮЂЩњОГжЎМфЕФЙиЯЕЁЃ
ЁЁЁЁЛиЕНЮЂЩњОГПижЦвђЫиЕФЬтФПЩЯРД‚ЪГЮяЙЉгІКЭбѕХЈЖШЪЧЮЂЩњОГЕФСНДѓПижЦвђЫи(ЭМ5)ЁЃдкШБгЊбјКЭИЛгЊбјЕФЛЗОГРя‚жЛгаЭтЩњжжКЭЧГФкЩњжжФмЙЛЩњДц‚вђЮЊдкШБгЊбјЕФЛЗОГРя‚ГСНЕЯТРДЕФгаЛњЮяДѓВПЗждкГСЛ§ЮяБэУцБЛЯћКФЁЂбѕЛЏ‚НігаЩйСПФбШмЕФгаЛњЮяОЙ§ЩњЮяШХЖЏЕШзїгУФмНјШыЧГФкЩњОГРя‚ЪГЮяЙЉгІЯожЦзХгаПзГцЕФЩњДцКЭЗБжГ;ЖјдкИЛгЊбјЛЗОГРя‚ бѕХЈЖШЪЧЯожЦвђзг(бѕКЌСПЕЭгк0.5mL/L ЪБ[18])‚ ДѓСПЕФгаЛњЮяБЛЪфЫЭЕНКЃЕз‚ЯћКФДѓСПШмНтбѕ‚ЩюЕФФкЩњОГвђШБбѕЖјЮоЩюФкЩњжжЩњДцЁЃжЛгадкжаЕШгЊбјЛЗОГРя‚ИїжжЮЂЩњОГЕФгаПзГцВХФмЙВЭЌЩњДц‚вђЮЊГ§СЫВПЗжгаЛњЮяБЛЭтЩњЕзЦмгаПзГцЯћКФЁЂбѕЛЏЭт‚ЛЙгаВПЗжгаЛњЮяжЪНјШыЩюЕФФкЩњОГРя‚дкетжжЧщПіЯТ‚ЪГЮяЙЉгІКЭбѕХЈЖШЙВЭЌЯожЦзХЕзЦмгаПзГцдкГСЛ§ЮяРяЕФзюЩюЗжВМЁЃЪГЮяЙЉгІЬЋЩйЛђепЬЋЖр‚ЖдКмЖрЕзЦмгаПзГцЖМВЛРћ‚ЪГЮяЙ§гкЦЖЗІЛсЕМжТгЊбјЙЉгІВЛзу‚ Й§гкЗсИЛЛсЕМжТГСЛ§ЮяДІгкЛЙдЛЗОГ‚етСНжжМЋЖЫОљЕМжТЕзЦмгаПзГцвдЬиЪтЛђЕЅвЛЕФЭтЩњжжЮЊжї‚ЩѕжСЕМжТЕзЦмгаПзГцЯћЪЇЁЃ
ЁЁЁЁ4 еЙЭћаТЪРМЭЩюКЃЕзЦмгаПзГцбаОП
ЁЁЁЁзнЙлЩюКЃЕзЦмгаПзГцбаОП‚ПЩвдЗЂЯж‚КЃЕзгаЛњЬМЭЈСПКЭЩюВПЫЎЭХетСНДѓвђЫиПижЦзХЕзЦмгаПзГцЕФЗжВМЁЃШчЙћЫЕЕзЦмгаПзГцдкКЃЕзГСЛ§ЮяРяЕФДЙЯђЗжВМЪмгЊбјЙЉгІКЭбѕХЈЖШЕФПижЦ‚ФЧУД‚ЦфдкбѓЕзКсЯђЩЯЕФЗжВМдђЪмЕБЕиЫЎЭХЫљПижЦЁЃКЃЫЎГСЛ§ЮяКЭКЃЫЎНчУцДІКЭНчУцвдЯТЕФГСЛ§ЮяРяЕФбѕХЈЖШ‚МШКЭгЊбјЙЉгІгаЙи‚гжКЭЫЎЭХБОЩэЕФбѕХЈЖШгаЙиЁЃМйШчКЃЕзгаЛњЬМЭЈСПВЛБф‚БЛаджЪВЛЭЌЕФЫЎЭХЫљИВИЧЕФКЃЕзЮЂЩњОГЕФ“жїШЫ”вВЛсКмВЛЯрЭЌЁЃвђДЫ‚ЮвУЧбаОПЕзЦмгаПзГц‚змЕФРДЫЕжївЊПЩзіСНМўЛљБОЗНУцЕФЪТЧщЃКвЛЪЧРћгУЕзЦмгаПзГцбаОПЩюВПЫЎЭХЕФБфЛЏЙцТЩ‚ЖўЪЧРћгУЕзЦмгаПзГцбаОПКЃбѓБэВуГѕМЖЩњВњСІЁЃЧАепЗДгГЕиЧђБэВуЯЕЭГжаЕФЫЎбЛЗ‚КѓепЩцМАЬМбЛЗ‚ЖМЪЧЕиЧђЯЕЭГПЦбЇжаЕФЛљБОЮЪЬтЁЃ
ЁЁЁЁНјШыаТЪРМЭ‚ДЋЭГЕФУшЪіадЕФЮЂЬхЙХЩњЮябЇвбОзЊШыаТвЛДњЕФЙХЛЗОГбаОП‚ВЛЭЌжЎДІдкгкЖЈСПЗжЮіЕФЗНЗЈЁЂзЗЫїЛњРэЕФФПБъКЭЖрбЇПЦЕФЭООЖЁЃзїЮЊЩюКЃЕзЦмЩњЮяжазюЮЊГЃМћЁЂгжФмБЃСєЛЏЪЏЕФУХРр‚ЕзЦмгаПзГце§ГЩЮЊЩюКЃЙХКЃбѓбЇбаОПЕФ“жїНЧ”‚еЙЯжзХ“ЕкЖўИіДКЬь”ЕФЧАОАЁЃгыДЫЯргІ‚баОПЗНЗЈЁЂЙлВтММЪѕЕФДДаТвбОГЩЮЊЪБДњЕФвЊЧѓ‚ШчЮЊЧѓЕУИќЭъећЕФЩњЬЌжИБъ‚дкРфВиЪвжаФЃФтКЃЕзЮТЖШЫЧбјЩюКЃЕзЦмгаПзГцЁЂВЩгУгЋЙтЕШаТММЪѕЪЖБ№ЛюгаПзГцЕШ[21]‚ ЖМЪЧгавцЕФГЂЪдЁЃ
ЁЁЁЁЯжГЁЕФЪЕЪБЙлВт‚НЋЪЧаТЪРМЭЩюКЃбаОПЕФживЊаТЗНЯђ‚ЕзЦмгаПзГцЮЊЬНВтЩюКЃКЃЕзЕФЙ§ГЬЬсЙЉСЫСщУєЕФМьВтЦїЁЃШчФЋЮїИчЭхББВП500ЁЋ600m ЫЎЩюДІЬўРраЙГіПкЗЂЯжЕФЕзЦмгаПзГц‚ЫЕУїдкзмЬхДІгкЛЙдЛЗОГЯТЕФКЃЕзЛЙПЩвдгаЖЬднМфаЊШУгаПзГцЩњДц[22]ЁЃЯёШеБОЯрФЃЭхЪНЕФЖЈЕуЙлВь‚ДгКЃУцЕНКЃЕзЕФЯЕЭГбаОП‚ЪЦБидкаТЪРМЭЕФЩюКЃбаОПжаНјвЛВНЭЦЙу‚ГЩЮЊНвЪОКЃбѓЙ§ГЬЕФаТЪжЖЮЁЃ
ЁЁЁЁЕБЧАЕФЙХКЃбѓбЇбаОП‚змЬхЦЋжиКЃЫЎЕФЩЯВуНсЙЙ‚ЖдгкКЃбѓЩюВПЕФаХЯЂУїЯдШБЗІЁЃЕзЦмгаПзГцЪЧЮЈвЛДгКЃАЖЕНЩюКЃХшЕиЮоЫљВЛдкЖјгжДѓСПГіЯжЁЂПЩвдБЃДцЕФЩњЮяУХРр‚ЪЧНЋЙХКЃбѓбЇЭЦНјЕНећИіКЃЫЎЯЕЭГЕФживЊжЇГХЁЃЭЈЙ§ВЛЭЌЩюЫЎКЃЕзЕФЕзЦмгаПзГцШЁбљбаОП‚ПЩЭћШЁЕУжаВуЫЎЁЂЩюВуЫЎКЭЕзВуЫЎЕФШЋЬзаХЯЂ‚БиНЋгажњгкДгИљБОЩЯМгЩюЖдЕиЧђЯЕЭГЕФРэНтЁЃ
ЁЁЁЁЕзЦмгаПзГцЕФИЦжЪПЧЬх‚ЙуЗКгУгкИїжжЮШЖЈЭЌЮЛЫи(Шчδ13C‚δ18O ЕШ)КЭдЊЫиБШжЕ(Шч Cd/Ca‚Mg/ Ca‚Sr/Ca)ЗжЮі‚вбОГЩЮЊЙХКЃбѓбЇЕФЛљБОЗНЗЈжЎвЛЁЃПЩЯЇЖдетаЉгаПзГцЕФЩњЬЌЬиеїСЫНтгаЯо‚вджСгкЖдЦфЙХЛЗОГНтЪЭЛљБОЩЯЪЧЕЅДПЕФПѓЮяЛЏбЇЗжЮі‚ ВЂВЛПМТЧПижЦетРрПЧЬхЩњГЄЕФгаПзГцЩњЬЌбЇЬиеїЁЃвђДЫаТЪРМЭЕФвЛДѓШЮЮёЪЧРЉеЙЩюКЃгаПзГцЕФЩњЬЌбаОП‚жЛгадкСЫНтЦфЮЂЩњОГЕФЬиеїКѓ‚ВХПЩФме§ШЗНтЪЭЦфПЧЬхЕФЛЏбЇГЩЗж[23]ЁЃ
ЁЁЁЁКЭЦфЫќУХРрЕФЛЏЪЏвЛбљ‚ЕзЦмгаПзГцЕФПЧЬхвВОРњзХТёВибЇЕФЩИбЁЃКНКНсПЧЕФЗжНтКЭИЦжЪПЧЕФШмНт‚ЕМжТТёВиШКЁЂЛЏЪЏШКгыЩњЮяШКжЎМфЕФВювьЁЃвђДЫ‚ТёВибЇЕФбаОПЪЧИљОнЛЏЪЏШКдйдьЙХЩњЬЌЛЗОГЕФБивЊЛЗНк‚МгЧПЕзЦмгаПзГцТёВибЇбаОП‚ЪЦБигаРћгкЙХЛЗОГбаОПЁЃ
ЁЁЁЁЮвЙњЕФЙХЩњЮябЇбаОПОпгаЙтШйЕФРњЪЗДЋЭГКЭГЄЦкЕФПЦбЇЛ§РлЁЃзїЮЊЙХКЃбѓбЇЁЂЙХЛЗОГбаОПЪжЖЮЕФЕзЦмгаПзГцбаОПЦ№ВННЯЭэ‚ЕЋНќ20ФъвбОШЁЕУСЫГЄзуЕФНјВНЁЃжаЙњзд1998ФъМгШыЙњМЪДѓбѓзъЬНвдРД‚ ЪзЯШЪЧвдЮЂЬхЙХЩњЮябЇЮЊЛљДЁЕФЙХКЃбѓбЇбаОПНјШыСЫЙњМЪбЇЪѕЧАби‚ПЊДДСЫЮвЙњЩюКЃбаОПЕФаТОжУцЁЃ 2003Фъ10дТ‚ЙњМЪзлКЯДѓбѓзъЬН(IODP)МЦЛЎЦєЖЏ‚ЮвЙњвдМгБЖЕФЭЖШыМЬајГЩЮЊЦфВЮгыГЩдБЙњ‚ЖјЙХЛЗОГбаОПЪЧ IODP ЕФШ§ДѓПЦбЇжїЬтжЎвЛ[24]‚вВЪЧЮвЙњПЩвдЗЂЛгзїгУЕФЪзбЁСьгђЁЃНќФъРД‚ЮвЙњЕФЩюКЃЕзЦмгаПзГцбаОПвбОзпЯђЙњМЪ(ШчЮФЯз25)‚дкаТЕФЙњМЪМЦЛЎжагІЕБНЋЪєжжзщКЯЕФбаОПгыЕиЧђЛЏбЇЕФЖржжжИБъЯрНсКЯ‚НЋЪвФкОЕЯТЗжЮігыКЃЩЯЯжГЁЙлВтЁЂЪдбщКЭЪ§жЕФЃФтЯрНсКЯ‚ељШЁдкИќИпЕФВуДЮЩЯВЮгыЙњМЪбЇЪѕКЯзїКЭОКељЁЃ——ТлЮФзїепЃКеХНгТ‚ЭєЦЗЯШ
SCISSCIAHCI