0
发表咨询在线!

发布时间:2020-01-20所属分类:农业论文浏览:1425次
摘 要: 摘要:黄瓜花叶病毒(Cucumbermosaicvirus,CMV)作为世界十大植物病毒之一,严重危害多种作物的生产。相对于药剂防治和使用转基因品种,利用植物自身的抗性基因培育抗病品种是最为经济有效的CMV防治方法。本文就相关内容从3个方面进行综述:(1)R基因介导的植物

摘要:黄瓜花叶病毒(Cucumbermosaicvirus,CMV)作为世界十大植物病毒之一,严重危害多种作物的生产。相对于药剂防治和使用转基因品种,利用植物自身的抗性基因培育抗病品种是最为经济有效的CMV防治方法。本文就相关内容从3个方面进行综述:(1)R基因介导的植物抗病毒机制及已克隆的R基因;(2)植物抗CMV的R基因及分子机制;(3)植物中其他抗CMV相关基因的研究。
关键词:植物;黄瓜花叶病毒;抗性基因
黄瓜花叶病毒(CMV)是雀麦花叶病毒科(Bromoviridae)黄瓜花叶病毒属(Cucumovirus)的典型成员,被列为世界十大植物病毒之一。黄瓜花叶病毒的寄主范围广,可侵染包括果树、蔬菜和观赏植物等100多个科的1000多种植物[1-2]。由于世界范围内有些地区存在多种病毒混合侵染等情况,CMV对作物生产的危害程度很难进行总体量化。根据科研人员的统计,在中国CMV的侵染可导致番茄减产25%~50%[3];在西班牙CMV的侵染可导致甜瓜减产60%,辣椒减产达80%[4-5];在西班牙一旦番茄上出现CMV侵染引发的坏死,产量损失达到80%以上,甚至绝收[6-7]。
蚜虫和种子带毒是CMV传播的主要途径,而药剂对病毒病的防治效果十分有限。利用基因工程技术进行抗CMV育种,具有周期短,不污染环境,抗性稳定等优点,但是转基因作物的安全问题仍受到多方质疑[8]。利用植物自身的抗性基因对植物进行遗传改良是最为经济和有效的遏制CMV危害的途径[9]。近几十年来,国内外学者对植物抗黄瓜花叶病毒基因进行了广泛和深入研究,并取得了一定的进展。本文就R基因介导的植物抗病毒病分子机制及已克隆的R基因植物抗CMV的机制和其他相关抗CMV基因的研究进展进行综述,以期为植物抗CMV基因的深入分析和应用研究提供参考。
1植物的天然免疫系统
一般认为植物免疫系统由2个层面的免疫反应组成:(1)植物通过细胞表面的跨膜识别受体识别病原物相关分子(Pathogen-associatedmolecularpatterns,PAMPs)所产生的免疫,称为病原物相关分子激发的免疫(PAMP-triggeredimmunity,PTI);(2)植物体内的抗性基因(Rgene)通过特异地识别病原物效应因子所产生的细胞内免疫反应,称为效应因子激发的免疫(Effector-triggeredimmunity,ETI)[10-11]。
1.1R基因介导的植物抗病毒机制
R基因介导的抗性是指R基因直接或间接识别特定病原物的无毒基因,激活植物防御反应。R基因介导的抗性可分为2种,一种称为系统获得性抗性(Systemacquiredresistance,SAR),主要表现为在病毒侵染点及周边组织引起程序性细胞死亡,活性氧爆发,细胞壁加厚,蛋白质磷酸化和去磷酸化以及大量防御基因的激活,随后整株植物出现非专一性的、抗多种与起始侵染病毒相关或不相关的植物病原的抗性。在SAR过程中,病程相关(PR)基因等防卫基因大量表达,信号转导因子水杨酸、茉莉酸、乙烯、一氧化氮等在植物抗病中起重要作用[11]。R基因介导的第2种抗性称之为极端抗性(Extremeresistance,ER)或细胞水平抗性(Cellularresistance),主要表现为有些R基因能够快速响应病毒的侵入,抑制病毒的积累,将病毒的侵染限制在单细胞水平,植物表型症状为侵染部位仅有极小的坏死点[12]。典型代表为马铃薯Rx1基因介导的对马铃薯X病毒的抗性以及番茄Tm-22基因介导的对烟草花叶病毒抗性,侵染部位不出现肉眼可见的超敏反应[13-14]。
1.2植物中克隆的抗病毒R基因
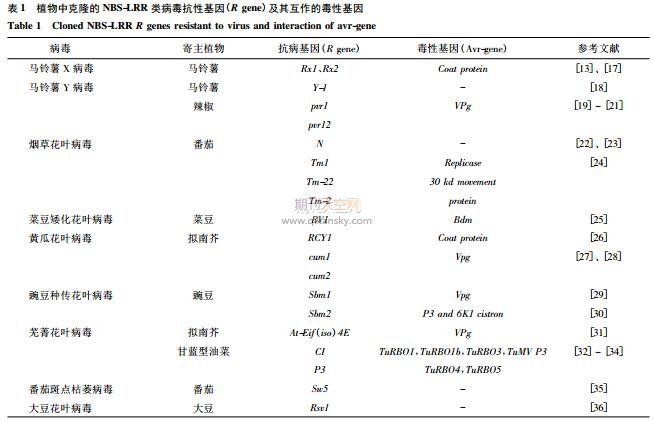
目前已有多种植物病毒的抗性基因被克隆[15-16],包括十几个典型的NBS-LRR类病毒抗性基因,其中N基因和抗马铃薯Y病毒(PotatovirusY,PVY)基因Y-1为TIR-NBS-LRR类病毒抗性基因,其余均为CC-NBS-LRR类病毒抗性基因(表1)。此外,还克隆了几个不属于NBS-LRR类病毒抗性基因家族的病毒抗性基因,包括拟南芥抗烟草蚀纹病毒(Tobaccoetchvirus,TEV)基因RTM1、RTM2和RTM3[37-38],番茄黄化曲叶病毒(Tomatoyellowleafcurlvirus,TYLCV)抗性基因Ty-1和Ty-3,番茄抗番茄花叶病毒(Tomatomosicavirus,ToMV)基因Tm-1[39]。
2植物中R基因介导的对CMV的抗性
2.1拟南芥中克隆的抗CMV的R基因及其抗性机制
拟南芥作为典型的模式植物,其CMV抗性基因克隆和机理研究最为广泛和深入,典型代表是抗CMV的单显性基因RCY1的研究。RCY1基因定位在5号染色体,可编码一个相对分子量为1.04×105的CC-NBS-LRR蛋白质,与抗霜霉病的RPP8基因和抗芜菁皱缩病毒的HRT基因为等位基因,抗性的发挥依赖于水杨酸(SA)和乙烯(ET)信号转导通路,不依赖于茉莉酸(JA)信号转导通路[26]。进一步研究发现SA和JA在调控RCY1功能方面存在拮抗作用,EDS5基因的表达和SA的积累在RCY1基因介导的抗性中发挥着重要作用[40-41]。RCY1基因表达量上调可显著增强植株对CMV的抗性,抗性方式表现为“基因对基因”模式[42]。RCY1通过LRR区域与CMV的致病因子CP蛋白互作诱导抗性[43]。拟南芥中WARKY70通过与RCY1基因编码的CC-NBS结构域互作,抑制CMV病毒的复制,减轻CMV的危害[44]。
推荐阅读:哪些农业专业可以申请职称
拟南芥中除典型的显性抗CMV的R基因RCY1,还克隆了2个典型的隐性抗CMV的R基因cum1和cum2,对应的CMV病毒致病因子为Vpg蛋白质。cum1和cum2可编码翻译起始因子4E(eIF4E)和4G(eIF4G)蛋白质,eIF4E蛋白质可与真核mRNA5’帽子结构结合,eIF4G属于支架蛋白质,能将eIF4E和起始因子结合形成eIF4F复合体,进而起始蛋白质合成。突变后的eIF4E不能和病毒移动蛋白质的mRNA结合,从而影响移动蛋白质的生成,限制病毒的移动[28]。
2.2其他植物中抗CMV的R基因及抗性机制
除拟南芥外,在其他植物主要是经济作物的抗CMV基因定位和克隆方面也取得了较多进展。菜豆中同源克隆的RT4-4基因编码一个TIR-NBS-LRR蛋白质,与CMV的2a蛋白质互作,引发过敏性坏死。该基因对番茄和辣椒上分离出的7个CMV株系具有抗性,但是对菜豆上分离的CMV株系无效[45]。2009年西班牙农业研究中心在甜瓜PI161375中鉴定出一个抗CMV基因cmv1[46],2017年该研究中心克隆了cmv1基因,该基因可以编码液泡分拣蛋白41,抗感材料间仅存在一个氨基酸差异(L348R),是目前唯一与CMV在韧皮部运输密切相关的隐性基因[47]。韩国科研人员在C.annuumcv.bukang中鉴定的抗CMV单显性基因Cmr1位于2号染色体的着丝粒位置,与番茄抗番茄花叶病毒的Tm-1基因和番茄抗CMV的QTL(QRCMV2)具有同位性,亚细胞定位结果显示Cmr1基因抑制了CMV病毒由叶表细胞到叶肉细胞的运动[48]。中国科研人员在C.frutescenscv.PBC688的2号染色体上定位到1个抗CMV主效QTL(qCmr2.1),预测的候选基因与拟南芥中抗CMV基因RCY1同属NBS-LRR家族基因,同时与烟草和番茄抗烟草花叶病毒基因N的同源性高达80%以上[49]。番茄抗CMV的QTL定位中发现2号染色体上的QTLQRCMV2与番茄抗番茄花叶病毒的Tm-1基因、辣椒抗CMV的Cmr1基因具有同位性[48,50-51];8号染色体上的QTL(QRCMV8)与马铃薯抗马铃薯S病毒的Ns基因具有同位性[51-52]。
3植物中其他相关基因介导的对CMV的抗性
植物通过多种基因协同作用形成的复杂调控网络抵御病毒侵害。R基因与致病因子的互作触发防御反应,多种基因的共同作用使植物获得系统抗性。所以多方位研究相关基因的抗性机制,是对植物抗CMV分子机制研究的深入和完善。
3.1拟南芥中其他相关基因介导的对CMV的抗性
拟南芥中除抗CMV的R基因外,防卫基因、转录因子、信号因子、光受体等也参与对CMV的抗性防御。①防卫基因:AGO1蛋白是拟南芥中RNA诱导沉默复合体中的核心组件,是抵抗CMV侵染的关键因子。CMV中2b蛋白可以阻断AGO1的酶切活性,限制miRNA途径,减弱RNA沉默效应,从而达到侵染的目的[53],拟南芥中过氧化氢酶基因CAT3与CMV的2b蛋白互作导致坏死斑的出现,CAT3基因的表达可增强拟南芥对CMV的抗性[54]。CMV的Fny株系的2b蛋白可以调控拟南芥中MircroR-NA159的表达水平诱发病症[55]。②转录因子:拟南芥中转录因子HAT1在抵抗CMV防御反应中起负调控作用,其表达依赖于SA的积累[56]。③信号因子:拟南芥中NbbZIP28为CMV病毒侵染应激的UPR信号调控因子,但不是唯一调控因子,在病毒侵染早期,可提髙寄主基础防卫反应,延缓病毒的侵染[57]。一氧化氮(NO)作为信号分子参与油菜素内脂介导的拟南芥对CMV的抗性反应[58]。④光受体:拟南芥光敏色素PHYB、向光素PHOT2基因显著影响抗性相关基因的表达和抗氧化剂的活性,在抵抗CMV侵染中起着重要作用[59]。
3.2其他植物抗CMV的相关基因
在烟草、油菜和苋色藜等植物中发现多种防卫基因参与植物对CMV的抗性。烟草锌指结构蛋白Tsip1与CMV1a和CMV2a蛋白结合形成复合体,抑制CMV的增殖[60]。烟草光敏色素信号通路通过调控内源SA信号通路发挥对CMV的抗性[61]。本氏烟热激蛋白NbHsp70与CMV病毒复制酶1a相互作用,促进CMV的复制而有利于病毒的侵染[62]。油菜中BnSGS3基因超表达可以抑制CMV的病毒积累,减轻CMV危害[63]。苋色藜CaNDR1a和CaNDR1b的转基因烟草对CMV的抗性增强,说明该基因参与了植物对病毒的内源免疫反应[64]。苋色藜CaNHO1基因可编码一种甘油激酶,其基因表达量在接种CMV后显著上调;转CaNHO1基因烟草可显著延缓CMV的晚期侵染,说明该基因参与了CMV诱导的防御抗性[65]。矮牵牛中转录因子PhERF2的表达可显著抑制CMV外壳蛋白基因的表达水平,水杨酸和乙烯可显著诱导该基因的表达,说明该基因可能是通过水杨酸和乙烯信号途径发挥作用[66]。
4讨论
综上所述,植物中已克隆了多个病毒抗性的R基因,为植物抗病毒病的深入分析和应用研究奠定了良好的基础。拟南芥抗CMV基因RCY1的研究为其他作物抗CMV基因的研究提供了较好的思路。但是由于技术局限性,从RCY1基因定位到分子机理的明确,整个研究历程超过10年。此外,由于植物对CMV的抗性大多是数量性状,其抗性机理更加复杂,研究历程更加漫长。例如茄果类蔬菜(主要是辣椒和番茄),自1997年Caranta等[67]在C.annuumcv.Perennial中首次定位到3个抑制病毒入侵的QTL以来,辣椒和番茄上仍未有抗CMV相关基因的克隆,其抗性机理更是模糊不清[68-69]。因此目前植物中克隆到的抗CMV的R基因仍然十分有限,抗CMV的分子机理仍需深入研究。克隆新的抗CMV的R基因并阐明其分子机理仍然是植物抗CMV研究的重点,也是作物抗CMV育种的技术和理论基础。
随着科技的发展,技术的不断革新,植物生物学研究方面的技术更加多样化,效率也在不断提高。首先,随着测序技术的发展,多种作物的基因组数据陆续公布。基于基因组数据产生的新基因定位技术(例如QTL-Seq、SLAF-Seq等)明显提高了基因定位效率,基于基因组数据开发出的新一代高密度分子标记(例如SNP和Indel等)使基因定位更为准确,这些为深入研究基因功能和抗性机理奠定了很好的基础[70-72]。在植物抗CMV基因定位方面,新技术的应用极为广泛,仅从受CMV危害最为严重的茄科作物辣椒来看,自2014年辣椒基因组数据公布以来,利用高通量测序的方法,新定位到10个抗CMV相关QTL,相对于之前的定位研究,耗费时间更短,定位区间更为准确[73-76]。其次,CRISPR/Cas9技术引领了分子生物学研究领域中颠覆性的技术革新。2013年5个独立的研究团队证明CRISPR/Cas9系统在真核生物中具有功能[77-81],最为重要的是研究证明该系统可以在多个位点同时实现高效的基因编辑[80-81]。由于CRISPR/Cas9技术具有简单、高效和易操作等优点,CRISPR/Cas9在基因功能研究和种质创制上具有巨大优势,在多种作物上已成功应用。例如在番茄上成功运用该技术编辑番茄早花基因SP5G,创制出可提早2周开花和果实成熟的番茄材料[82];美国冷泉港实验室的科研人员利用该技术编辑SICLV3基因启动子序列创制出系列番茄产量相关性状的突变体材料,实现了对数量性状更加微小的调控[83]。虽然目前还未有利用CRISPR/Cas9技术研究植物抗CMV分子机理的报道,但是利用该技术进行抗CMV分子机理研究是一种可行的思路,也是目前创制突变体材料及抗性种质材料最为高效的途径。所以充分利用多种高效的生物技术实现植物抗CMV基因研究的突破,将是科研人员进一步研究和探索的重点。
婵犵數濮伴崹鐟帮耿鏉堛劍娅犳俊銈傚亾閸楅亶鏌ㄩ悤鍌涘:闂傚倷绀侀崯鎸庢櫠鎼淬垺鍙忛柣銏⑶归崙鐘绘煕閹伴潧鏋熼柛瀣墬閵囧嫰寮崶顬捇姊婚崟顐ばч柡灞诲€栫缓鑺ュ緞婢跺本鏅奸梻浣虹帛閹碱偆妲愰弴鐘愁潟闁哄啫鐗嗙粻锝嗙箾閸℃瀚板ù婊勫劤闇夐柨婵嗘搐閸斿淇婇姘捐含闁哄矉绻濆畷顏呮媴閸涘﹦浜栭梻渚€鈧偛鑻崢鍛婁繆閻愭潙绗х紒顔规櫆閵堬綁宕橀妸锔绘闂備線娼荤€靛矂宕㈡ィ鍐ㄧ闁稿繘妫跨换鍡樸亜閹邦喖鏋庡ù婊冨⒔缁辨挻鎷呮搴樻晙闂佹悶鍔嶅钘夘嚕閹惰姤鍋勯柣鎾虫捣椤斿顪冮妶鍡橆梿闁稿鍔曢~婵嬪Ω閳哄倻鍙嗗┑鐐村灦宀e潡鎮為柨瀣ㄤ簻闁哄洦顨嗗▍濠勨偓娈垮枤婢ф骞嗛弮鍫濐潊闁斥晛鍟浼存⒒娴e憡鎯堥柛濠勄圭叅闁靛ě鍌滃墾闂佺粯鍔曞Ο濠囧疮閸涘瓨鐓犵痪鏉垮船婢т即鏌i妷銉﹀殗闁哄本绋戦埢搴ㄥ箛椤掑倷绱橀梻渚€娼уΛ妤呭疮娴兼潙绠熼悗娑櫳戞刊瀵哥磼鐎n厽纭跺ù婊勭墵濮婃椽骞愭惔锝傛闂佸搫鐗滈崜姘扁偓闈涖偢瀹曟﹢顢欓懖鈺冨炊婵犵數濮磋墝闁稿鎸剧槐鎾愁吋閸℃﹩妫冮梺纭呮珪閹瑰洭寮幘缁樻櫢闁跨噦鎷�.闂傚倷绀侀崯鍧楁倶閸儱鐒垫い鎺嶇婢ь垱绻涚仦鍌氣偓婵嬪蓟閿涘嫧鍋撻敐搴′簽闁活厼顑夐弻娑氣偓锝庝簽鏁堥悗娈垮枙閸楁娊銆佸☉妯锋瀻闁瑰濮弸娆撴⒒娴gǹ鎮戞繝銏★耿楠炲﹤顓奸崶鈺冪暥闂佸搫绉查崝搴ㄥ煝閺冨倵鍋撻獮鍨姎婵☆偅顨婂畷銏ゅ箳濡や胶鍘搁梺鍓插亽閸嬪嫭鏅堕鈧铏规喆閿濆棙鐝氶悗瑙勬穿缂嶄礁顕i崜浣瑰磯闁靛⿵濡囩粔鍓х磽閸屾瑨顔夐柛瀣崌閺岋綁骞嬮悙鍡樺灴閹顢氶埀顒勫蓟閿濆拋娼╅柣鎾冲閻忓棙绻涢敐鍛悙闁挎洦浜悰顔界瑹閳ь剟寮幘缁樻櫢闁跨噦鎷�,闂傚倷绀侀幉锟犳偡椤栫偛鍨傞柛顭戝亞椤╃兘鏌涢鐘茬仼閻庡灚鐓¢弻銊╂偆閸屾稑顏�闂備浇顕х€涒晠宕樻繝姘挃闁告洦鍋撻懓鍧楁煣韫囷絽浜栧ù婊勭矊铻栭柨婵嗘噹閺嗙偤鏌ㄥ☉铏婵犵數鍋涢悺銊у垝瀹€鍕亱闁告侗鍠氶々鏌ユ煙閻戞﹩娈旂紒鈧崱娑欑厽婵☆垵鍋愮敮娑㈡煟濠垫挻瀚�.
闂傚倷绀侀幉锟犲吹鎼淬劌绀夐煫鍥ㄤ緱閺佸淇婇妶鍛伀闁汇倐鍋撻梻浣告啞閸擃剟宕橀妸褝绱甸梻鍌氬€风欢锟犲磻閸曨垁鍥偨閹肩偐鍋撻崒鐐存櫢闁跨噦鎷�

SCISSCIAHCI

