0
发表咨询在线!

发布时间:2022-01-10所属分类:园林工程师浏览:1653次
摘 要: 摘 要: 自然条件下晴天银杏叶片光系统Ⅱ 光化学效率 ( Fv /Fm)表现明显日变化。 上午 Fv / Fm随光照的增强而降低 ,至 14: 00左右达最低值。其后随着光强的减弱 Fv / Fm缓慢恢复。一天中叶黄素循环关键组分玉米黄质 ( Z)含量与 Fv / Fm呈负相关。用二硫苏糖醇 ( DTT)阻

摘 要: 自然条件下晴天银杏叶片光系统Ⅱ 光化学效率 ( Fv /Fm)表现明显日变化。 上午 Fv / Fm随光照的增强而降低 ,至 14: 00左右达最低值。其后随着光强的减弱 Fv / Fm缓慢恢复。一天中叶黄素循环关键组分玉米黄质 ( Z)含量与 Fv / Fm呈负相关。用二硫苏糖醇 ( DTT)阻断 Z 的形成后 ,光抑制程度大大加深。 结果表明与叶黄素循环有关的非辐射能量耗散的增加是产生光抑制的原因之一。强光处理前饲喂 D1蛋白合成抑制剂林可霉素 ( LM) , Fv /Fm下降幅度增加 ,而且在暗中与弱光下的恢复出现差异 ,但强光处理后再引入 LM,则对 Fv /Fm 两种恢复无明显影响。 这表明强光可引起银杏叶片 D1蛋白的快速周转而植物体内这种 D1蛋白钝化、复活之间的动态平衡可能是避免强光损伤的保护性反应。 自然条件下强光胁迫并不能导致银杏叶片 D1蛋白的净损失。
关键词: 光抑制; PSⅡ 光化学效率;叶黄素循环 ; D1蛋白;银杏
强光胁迫下植物光合作用的光抑制受到越来越多研究者的关注。 尤其是近年来随着天然色素非辐射能量耗散过程的发现 ,人们对光抑制机理的认识已发生了很大的改变 (许大全 1992, Long& Humphries 1994)。 我们曾观测到晴天中午银杏叶片有严重光合午休现象 ,并推测与光抑制有关 (孟庆伟 1995) ,但一天中 Fv /Fm下降程度和叶黄素循环各组分含量的关系 ,特别是和玉米黄质 ( Z)的关系目前还不清楚。关于 D1蛋白周转 ( turnov er) 与光抑制的关系一直众说纷纭。 Ohad( 1984)认为光系统Ⅱ 反应中心的钝化与 D1蛋白有关。 D1蛋白周转发生在光抑制过程的早期阶段 ,而 Clela nd( 1990)则认为光系统Ⅱ 失活发生在 D1蛋白净损失之前 , D1蛋白降解是为了阻止光系统Ⅱ反应中心失活程度的进一步加深。郭连旺 ( 1994, 1996)对自然条件下珊瑚树及田间小麦的观测并未发现 D1蛋白的净损失。本文将研究银杏叶片光合作用光抑制的机制 ,着重探讨叶黄素循环及 D1蛋白的周转与光抑制的关系。
1 材料与方法
1. 1 材料栽培
两年生银杏苗自然条件下室外盆栽。盆内径 45 cm,盆高 60 cm,装干土 16. 5 kg ,常规管理。
1. 2 叶绿素荧光参数的测定
采用便携式植物效率分析仪 (英国 Ha nsa tech公司制造 )测定初始荧光 ( Fo) ,最大荧光 ( Fm)和光系统Ⅱ光化学效率 ( Fv /Fm)。 测前预暗 5 min,闪光时间 2 s。
1. 3 叶黄素循环各组分分析
用直径 1 cm 的打孔器取叶圆片 8~ 10片 ,放入研钵 ,加少许石英砂与 CaCO3 ,用 100% 丙酮研成匀浆。 1000 r /min离心 5 min。 叶黄素循环各组分的高压液相色谱分析参照赵世杰 ( 1995)方法。 叶绿素 a( chl a)和叶绿素 b( chl b)含量用 UV -120分光光度计测定。
1. 4 叶片气体交换参数的测定
光合速率 ( Pn)及光量子通量密度 ( PFD)由便携式光合气体分析系统 (英国 ADC公司制造 )获得。 图中所列数据为 8~ 10片叶的平均值。
1. 5 抑制剂的引入
将选好叶片的叶柄于水下再剪一段 ,迅速转移至 30 mmo l /L的二硫苏糖醇 ( DT T)或 3 mmo l /L的林可霉素 ( LM)溶液中 ,分别在弱光 (约 30μmol· m - 2· s - 1 )条件下放置 10 或 3 h。 对照叶片浸于蒸馏水中。
2 结 果
2. 1 银杏叶片光合速率及叶绿素荧光参数日变化
从图 1可以看出 ,银杏叶片光合速率 ( Pn)日变化曲线为双峰型。 Pn第一次高峰出现在上午 10: 00左右。此时光强 ( PFD)约为 1200μmo l· m - 2 · s - 1 。下午 14: 00前后 , Pn达最低值 (光强约 1600μmo l· m - 2· s - 1 )。此后 Pn略有回升 ,至 16: 00达第二次高峰 ,然后随着光强的减弱而降低。
光系统Ⅱ 光化学效率 ( Fv /Fm)则随光照的增强而降低。低谷出现在 14: 00左右 ,与早晨 ( 6: 00)相比 ,下降 21. 8%。至下午 18: 00恢复 90%以上。最大荧光 Fm也表现相似的变化趋势。与此相反 , Fo先升高 ,后降低 ,但变化幅度不大 (图 2)。 Fv /Fm的下降主要由 Fm 降低引起。
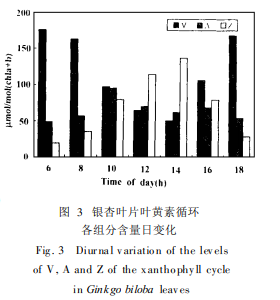
2. 2 叶黄素循环各组分含量日变化
图 3表示叶黄素循环三组分紫黄质 ( V)、环氧玉米黄质 ( A)和玉米黄质 ( Z)含量日变化。上午随着 PFD升高 ,光抑制加重 (图 2)。紫黄质 ( V )的脱环化作用也随之加强 , Z的含量增高。下午 Fv /Fm降至最低时 , Z的含量也达最高值。 其后随着光强减弱 , Z的含量下降。以上结果表明 ,与叶黄素循环有关的非辐射能量耗散的增加可能是光系统Ⅱ光化学效率下降的原因之一
2. 3 叶黄素循环抑制剂 D TT对银杏叶片光抑制的影响
二硫苏糖醇 ( DT T)引入叶片后 ,与对照叶片同时放在强光 ( 1600 μmol· m - 2· s - 1 ) 下处理 2 h ,再测定叶片的荧光参数及恢复过程 ,结果如图 4。与对照相比 , DTT处理的叶片 Fv /Fm下降幅度增加了 29% 。 表明抑制玉米黄质的形成后光抑制程度加重。 并且 DTT 处理的叶片 ,黑暗及弱光下恢复 ( 50 μmo l· m - 2· s - 1 )程度不同。 银杏叶片在暗处的不完全恢复 (恢复约 67% )表明抑制叶黄素循环后 ,可能引起 D1蛋白净降解。
2. 4 D1蛋白合成抑制剂 LM 对叶片光抑制及其恢复的影响
图 4 DTT处理对银杏叶片光抑制程度及恢复的影响 Fig . 4 Pho toinhibitio n a nd its r eco ve ries under DTT trea tment 为进一步探讨强光对银杏叶片 D1蛋白的影响 ,我们观察了 D1蛋白合成抑制剂林可霉素 ( LM )对叶片光抑制及其恢复过程的影响。 发现饲喂 LM 的叶片遭受 2 h 强光 ( 1600 μmo l· m - 2· s - 1 )照射后 , Fv /Fm的下降程度比对照增加了 39. 7% ,而且暗中放置 16 h 后 ,只恢复了 25% (图 5)。 表明此时强光造成了 D1蛋白的破坏。 但在恢复阶段引入 LM 暗恢复及弱恢复均可达 95%以上。 说明 2 h强光处理后并未发生 D1蛋白的净损失。
3 讨 论
自 50年代 Hag er发现强光照射的绿色植物叶片存在叶黄素含量的变化以来 ,叶黄素循环对光合器官的保护作用已得到多数研究者的认同 ,不仅紫黄质的脱环氧化能耗散过剩激发能 ,清除活性氧 ( Demmig-Adamas 1990, 1992) , Z本身也参与叶绿体色素复合物的能量耗散。 Z的含量与非光化学猝灭呈线性相关 ( Lo ng a nd Humphries)即是一有力证据。本实验观察到银杏叶片光合日进程中玉米黄质含量与 Fv /Fm下降幅度密切相关 (图 3) , 饲喂 DT T后 Fv /Fm下降程度增大 (图 4) ,光抑制加重 ,并导致 D1蛋白的损失 ,表明依赖叶黄素循环的非辐射能量耗散的增加是银杏叶片产生光抑制的原因之一。
D1蛋白周转一直是光抑制机理研究中心较活跃的领域 ,但因材料、方法及环境条件的不同 ,至今人们仍未达成共识。 我们通过比较强光处理前后引入 D1蛋白合成抑制 ( LM )光抑制恢复过程的差异 ,发现强光处理前引入 LM能造成 D1蛋白损失 (暗恢复不完全 ) ,而强光处理后引入 LM,暗恢复及弱光恢复无明显差异 (图 5) ,表明单纯的强光胁迫未引起 D1蛋白的净损失。 由此可知 ,强光胁迫下存在 D1蛋白的周转 ,但自然条件下 D1蛋白的钝化、复活达到动态平衡并不表明 D1蛋白的净降解。 根据以上分析 ,我们推测自然条件下银杏叶片叶黄素循环引起的非辐射能量耗散和 D1蛋白的可逆失活是导致光系统Ⅱ光化学效率下降的两个主要因素。——论文作者:张 宁 ,孟庆伟 ,赵世杰 ,许长城 ,邹 琦
参考文献:
[1 ] 许大全 ,张玉忠,张荣铣.植物光合作用的光抑制 [ J].植物生理学通讯 , 1992, 28( 4): 237~ 243.
[2 ] LON G S P, HUM PHRIES S. Pho toinhibiti on of phot osynthesis in nature [ J]. Annu. Rev. Plant. Physiol. Plant Mol. Biol. , 1994, 45: 633~ 662.
[3 ] 孟庆伟 ,王春霞,赵世杰等 .银杏光合特性的研究 [ J].林业科学 , 1995, 31( 1): 69~ 71.
[4 ] O HAD I, KYLE D J, ARN TZEA C J. M emb rane protein damage and repair, removal and replacement of inactivat ed 32-kil odalton polypeptides. in chlo ropl ast membranes[ J]. J. Cell. Biol. , 1984, 99: 481~ 485.
[5 ] CLEL AND R E, DEM MIG ADAMS B, ADAMS W W Ⅲ et al. Phos ph orylati on stat e of th e ligh tharvesting chl oroph yll prot ein complex of ph ot os yst emⅡ and chl orophyll fluorescence charact eristics in Monstera d eliciosa Liebm. and Glycine ma x ( L. ) [ J]. Merrill in res ponse to ligh t. Aust. J. Plant Physiol. , 1990, 17: 589~ 599.
[6 ] 郭连旺 ,许大全.自然条件下珊瑚树叶片光合作用的光抑制 [ J].植物生理学报 , 1994, 20( 1): 46~ 54.
[7 ] 郭连旺 ,许大全,沈允钢.田间小麦叶片光合作用的光抑制不伴随 D1蛋白的净降解 [ J].植物学报 , 1996, 38( 3): 196~ 202.
[8 ] 赵世杰 ,孟庆伟,许长城等 .植物组织中叶黄素循环组分的高效液相色谱分析法 [ J].植物生理学通讯 , 1995, 31 ( 6): 438~ 442.
[9 ] DEM MIG ADAM S B. Carot enoid and phot oprot ection in pl an ts: A role f or the Xanth oph yll Zeaxanthin [ J]. Biochimica et Biophysica Acta. , 1990, 1020: 1~ 24.
[10 ] DEM MIG ADAMS B, ADAM S W WⅢ . Phot oprot ection and oth er res ponses of plants to hi gh li ght stress [ J]. Annu. Rev. Plant Physiol. Plant Mol. Biol. , 1992, 43: 59~ 62
声明:①文献来自知网、维普、万方等检索数据库,说明本文献已经发表见刊,恭喜作者.②如果您是作者且不想本平台展示文献信息,可联系学术顾问予以删除.

SCISSCIAHCI

