0
发表咨询在线!

发布时间:2019-09-16所属分类:医学职称论文浏览:1124次
摘 要: 摘 要:机体每时每刻都有大量细胞凋亡,及时清除凋亡细胞对维持机体的免疫平衡至关重要,凋亡细胞被吞噬细胞清除过程分为:招募、识别和吞噬消化3个阶段。凋亡细胞清除表现为没有炎性反应或者一个已经存在的促炎反应在吞噬发生后被下调。当机体处于组织损伤

摘 要:机体每时每刻都有大量细胞凋亡,及时清除凋亡细胞对维持机体的免疫平衡至关重要,凋亡细胞被吞噬细胞清除过程分为:招募、识别和吞噬消化3个阶段。凋亡细胞清除表现为没有炎性反应或者一个已经存在的促炎反应在吞噬发生后被下调。当机体处于组织损伤和炎性反应状态或生理应激时,细胞发生凋亡或坏死,若凋亡细胞不能及时清除也会继发性坏死,细胞坏死释放内源性危险信号分子,也称为危险相关分子模式(DAMPs)。DAMPs在结构和功能上与侵入机体的外源性的保守的微生物即病原体相关分子模式的表面结构具有相似性,可被病原体识别受体(PRRs)识别,激活相关信号通路,产生自身抗体或活化其他免疫细胞,导致自身免疫性疾病或炎性反应。
关键词:细胞凋亡; 细胞坏死; 死亡细胞清除; 胞葬
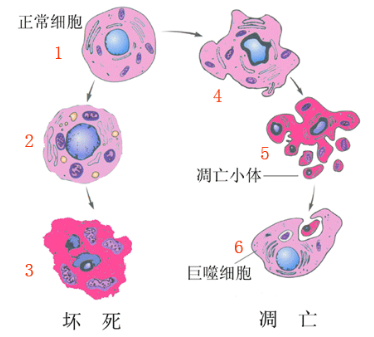
死亡是生命的一部分,多细胞生物的出生和生命维持每日需要数以亿计的细胞死亡贯穿其生命周期。所谓细胞死亡的定义包括3个方面:(1)永久失去细胞膜的完 整 性;(2)细胞 裂 解 为 碎 片(主 要 指 凋 亡 小体);(3)被职业和非职业吞噬细胞吞噬。19世纪后期LLYA METCHINKOFF 因 发 现 吞 噬 细 胞 (phago-cytes)于1908年获得诺贝尔生理医学奖。细 胞 死 亡是机体发育、应激、感染和自稳的重要组成部分,正常成人每一天有500亿个细胞死亡,细胞死亡后能够迅速被邻近细胞或巨噬细胞识别吞噬及消化,这个过程称为胞葬(efferocytosis)。死亡细胞自身或细胞死亡时释放的物质有利于死亡细胞的清除及免疫学转归。死亡细胞清除 缺 陷 与 自 身 免 疫 性 疾 病 和 自 身 炎 性 疾病密切相关,如系统性红斑狼疮、动脉粥样硬化、慢性阻塞性肺疾病和糖尿病等[1]。
1 细胞死亡的主要方式
生命是由单个细胞发育而来,它需要分化成不同形式的细胞,进而形成一个复杂的有机体。在发育和后续生命中,伴 随 着 数 亿 个 细 胞 的 死 亡,有 些 细 胞 死亡是生理性的,有些是病理性的。很多科学家描述了自然发生的细胞死亡,然而大部分生物学家是对细胞的 生 命 更 感 兴 趣。1965 年,LOCKSHIN 和 WIL-LIAMS首次提出程序化细胞死亡的概念,它 描 述 了细胞死亡是细胞在胚胎发育中,在特定空间和特定时间程序化死去的一种过程。1972年 KERR 根据这种在发育和组织稳态中发生的细胞死亡的形态学特征,提出了凋亡(apoptosis)一词(来 源 于 希 腊 字,意 思 是落下,好像 叶 子 从 树 上 落 下),将它 与 坏 死(necrosis)即一种由物理化学等因素导致的细胞死亡区分开来[2]。
这些年来,很多研究者对细胞死亡的形态特征、理化性质、其发生的机制以及对机体的影响展开了深入研究,发现了 更 多 不 同 类 型 的 细 胞 死 亡 方 式,从 不同角度予以 不 同 的 分 类。细 胞 死 亡 可 根 据 其 形 态 分类:细胞凋亡、坏 死、细 胞 自 噬 或 有 丝 分 裂 灾 难 等;可根据有无核酸酶或不同蛋白酶的酶学标准(半胱天冬酶,钙激活中性 蛋 白 酶,组 织 蛋 白 酶 和 谷 氨 酰 胺 转 氨酶)分类;还可 以 根 据 功 能(程 序 性 或 意 外,生 理 或 病理)分类;或根据免疫学特性(有或无免疫原性)分类。2009年和2012 年细 胞 死 亡 命 名 委 员 会(NCCD)[2-3]建议根据形态的标准统一分类细胞死亡,目前发现细胞死亡 有 凋 亡、坏 死、自 噬 性 细 胞 死 亡 (autophagiccelldeath)、角化 性 细 胞 死 亡(cornification)、非典 型细胞死亡。非典型细胞死亡又包括有丝分裂灾难、失巢凋亡(anoikis)、沃勒变性(walleriandegeneration)、副凋亡(paraptosis)、细胞焦亡(pyroptosis)、焦亡性细胞坏死(pyronecrosis)、坏死 性 凋 亡(necroptosis)、内亡(entosis)、兴奋性中毒(excitotoxicity)、胞外诱捕网致死(ETosis)、铁死 亡(ferroptosis)以及红细胞衰亡(suicidalerythrocytedeathoreryptosis)。2015年和2018年 NCCD 建 议 增 加 功 能 和 分 子 机 理 分 类[4-5]。2015年 NCCD将细胞死亡分为意外性细胞 死 亡(ac-cidentalcelldeath)和调节性细胞死亡(regulatedcelldeath),调节性细胞死亡又包括程序性细胞死亡(pro-grammmedcelldeath)。
2 胞葬的机制
吞噬就是摄入大小超过0.5μm 的颗粒。在这个过程中,巨噬细胞通过细胞骨架重排包围即将被消化的颗粒,形成 一 个 所 谓 的 吞 噬 环。环绕颗粒完成后,颗粒就被包埋 在 质 膜 小 囊 中,形 成 了 吞 噬 小 体,随 后它成熟为吞噬溶酶体。
吞噬细胞不总是邻近死亡细胞,所以它们必须被招募,被引诱 到 死 亡 细 胞 处 就 餐。职 业 吞 噬 细 胞(巨噬细胞和树 突 状 细 胞)和 非 职 业 吞 噬 细 胞(如 上 皮 细胞或内皮细胞)迁移到死亡细胞处,通 过 细 胞 表 面 受体识别和吞 噬 死 亡 细 胞,内 化(常 是 整 个 吞 噬),降解和处理。对于凋亡细胞此过程是免疫耐受的;胞葬过程常分为4步:(1)“找到我(find-me)”信号招募吞噬细胞;(2)“吃我(eat-me)”信号识别和束缚死亡细胞;(3)吞噬死亡 细 胞;(4)处理、降解和对吞噬残骸的免疫反应。
2.1 凋亡细胞清除过程的阴与阳
2.1.1 “找到我(find-me)”信号 吞噬细胞必须被凋亡细胞分泌的所谓的“找到我(find-me)”信号所趋化,S19核糖体蛋白二聚体(dRPS19)是第一个被发现的凋亡细胞趋化信号,它是核糖体的亚单位成分。发生交联的 RPS19可趋化单核细 胞 和 巨 噬 细 胞,其 原 因与 G 蛋白偶联受体 CD88相关。内皮-单核细胞激活多肽Ⅱ(EMAPⅡ)是从甲基胆蒽 A 条件培 养 基 培 养的小鼠成纤维肉瘤细胞中分离到的一种多肽,EMAP Ⅱ对中性粒细胞和单核细胞均有化学趋化性,并能诱导中性粒细胞 髓 过 氧 化 物 的 释 放 及 单 核 细 胞 肿 瘤 坏死因子(TNF)-α的产生[1,6]。
生发中心 B细胞在亲和力成熟过程中增加凋亡,这些凋亡细胞释放膜相关 CX3CL1(或fractalkine)微颗粒,被巨噬细 胞 经 典 趋 化 受 体 CX3CR1感知,介 导巨噬细胞到凋亡细胞处。
胱天蛋白酶活化磷脂酶 A2导致产生和释放溶血卵磷脂(LPC),LPC被 G 蛋白偶联受体 G2A 感知,刺激巨噬细胞招募。同样凋亡细胞释放的1-磷酸-鞘氨醇(S1P)被多 种 G 蛋 白 偶 联 受 体 S1P-R1-5识别,介导吞噬细胞 趋 化。然 而 这 些 磷 脂 在 体 内 循 环 中 的 浓度高于凋亡细胞释放的浓度,它们的体内功能没有很好评估。因此,这种趋化活性仅仅是局部的。
可能最有潜 力 的“找 到 我”信 号 是 核 苷 酸。通 过活化缝隙链接蛋白1通道(Panx1)以胱天蛋白酶依赖释放 ATP 和 UTP,被 吞 噬 细 胞 表 面 嘌 呤 受 体 如P2Y2探 知。体内用糖皮质激素治疗干扰核苷酸/P2Y2相互作 用 导 致 凋 亡 胸 腺 细 胞 增 加。ATP 既是 “找 到 我 ”信 号,同 时 也 是 危 险 相 关 分 子 模 式(DAMPs),可警 告 天 然 免 疫 系 统。坏死细胞释放的尿酸既是“找到我”信号,同时也是炎症小体的活化因子。
凋亡细胞释放的乳铁蛋白(lactoferrin)是已知的 “keep-out”信号 或 抗 趋 化 信 号(anti-chemotacticsig-nal),死亡中性粒细胞和嗜酸性粒细胞释放,对中性粒细胞抗趋化作用;乳铁蛋白可以选择性抑制嗜酸性粒细胞的募集。但 是 它 对 单 核 细 胞 和 巨 噬 细 胞 向 补 体成分 C5a的趋化却没有影响。活细胞表达的补体调节因子如 CD46和 H 因子抑制补体活化瀑布起到潜在抗 趋 化 作 用。总 之 吞 噬 细 胞 感 知 “找 到 我 (find-me)”信 号 和 “keep-out”信 号,两种在体内处于平衡状态。
2.1.2 “吃我(eat-me)”信号 细胞死亡的环境是复杂的,有多种细 胞 存 在,包 括 死 亡 细 胞、健 康 细 胞、免疫细胞。吞噬 细 胞 必 须 从 众 多 的 细 胞 中 区 分 出 死 亡细胞,这样才能 维 持 稳 态,促 进 正 常 的 发 育 并 避 免 不期望的炎症发生。
在凋亡的过程中新的分子会出现在细胞的表面。最熟知的就是磷脂酰丝氨酸(phosphatidylserine)从细胞膜的内 表 面 进 入 外 表 面。钙 离 子 介 导 的 阳 离 子通道 TMEM16F介导脂质颠倒(lipidscrambling),胱天蛋白 酶-3切割 拼 接 酶 Xkr8,完成磷脂酰丝氨酸外翻。翻转酶(flippase)ATP11C 正常 将 氨 基 磷 脂 从 胞外转移到胞浆,凋亡时胱天蛋白酶-3酶切 ATP11C使其失去活性。
尽管磷脂酰丝氨酸在很多生物膜是少量组分,但是磷脂酰丝 氨 酸 有 很 重 要 生 物 学 功 能。膜 外 暴 露 的磷脂酰丝氨酸是“吃我(eat-me)”信号,是凋亡细胞识别和清除的必需组分。吞噬细胞通过膜受体结合,如T 细胞免疫球蛋白域黏蛋白域受体-4(Tim-4)、脑特异血管生成抑制因子-1(BAI-1)、稳定素-2(stabilin-2)等。也可以通过体液中的桥梁分子结合,如乳脂球表皮生长因子-8(MFG-E8)、GAS-6、蛋白S,识别吞噬细胞表面 受 体 如 整 合 素 αvβ3、αvβ5或 TAM 受体(Ty-ro3-Axl-Mer)。磷脂 酰 丝 氨 酸 与 相 应 受 体 结 合 后 导致细胞骨架重排,完成吞噬细胞残骸[1,7-8]。
磷脂酰丝氨酸对巨噬细胞识别和吞噬的重要性首先发现于红细胞,衰老的红细胞清除跟凋亡细胞类似,依赖 Fas-半胱 天 冬 酶-3/半胱 天 冬 酶-8信号 导 致的磷脂酰丝氨酸外翻;还有 N-乙酰葡萄糖胺和岩藻糖的表达增加。这 些 变 化 累 积 成 一 个 对 吞 噬 细 胞 来 说代表强烈的“吃我(eat-me)”信号的凋亡特异性的表面模式。
正常情况下活细胞或活化细胞外膜磷脂酰丝氨酸浓度极低,不被吞噬。即使组成性表达 TMEM16F,也不被吞噬。吞 噬 细 胞 如 何 区 别 磷 脂 酰 丝 氨 酸 阳 性 的死亡细胞和活细胞呢?事实上活细胞表面表达“不吃我(don′teat-me)”信号,如 CD31、CD47和 CD61。活细胞表面表达 CD31、CD47和 CD61,阴性调节吞噬细胞吞 噬,即使这些细胞表达磷脂酰丝氨酸也不被吞噬,见图2。
活细胞上的 CD47,又称整联蛋白(IAP),与巨噬细胞上的信号调节蛋白α链(SIRPα)相互作用可以负相调控吞噬作 用,并对固有免疫系统产生抑制作用。如果存在于红细胞表面的 CD47表达减少,则脾的红髓区巨噬细胞对红细胞的吞噬作用增强,这也是溶血性贫 血 的 发 病 因 素 之 一;如果存在于血小板表面的CD47表达减少,则会导致巨噬细胞吞噬破坏血小板,这与特发性血小板减少性紫癜发病紧密相关。同 样活细胞上的 CD31可以与吞噬细胞上的 CD31相互作用,提供一个空间上的排斥作用来抑制吞噬作用。不同细胞 CD31表达不同,某些白细胞 CD31的表达下调对有效率的吞噬是必要的。
推荐阅读:细胞成像分析论文发表期刊
细胞成像分析在生物学或者医学中是经常用到的,很多这方面的研究人员也会发表这类的论文,但是他们对于这类论文可以投稿的期刊了解的不太清楚,为此期刊天空的小编查阅了相关资料,并且和以往评职人员进行分析,在这里给大家推荐了能接收这类论文的期刊,需要发表论文的作者可以作为参考,或者您可以直接咨询我们的编辑老师了解详情。
此外,磷脂酰丝氨酸不是唯一“吃我(eat-me)”信号。其他如ICAM-3,氧化 LDL-样分 子、糖 化 表 面 蛋白、C1q结合血清蛋白均被认为是“吃 我(eat-me)”信号。胞内内质网上的钙网蛋白(calreticulin)转运到胞浆外膜也作为“吃我(eat-me)”信号被吞噬细胞识别。胞葬被“吃我(eat-me)”信号和“不吃我(don′teat-me)信号”平衡调 节,很 清 楚 死 亡 细 胞 通 过 表 达 表 面 识 别分子促进自身被吞噬细胞识别和清除。
声明:①文献来自知网、维普、万方等检索数据库,说明本文献已经发表见刊,恭喜作者.②如果您是作者且不想本平台展示文献信息,可联系学术顾问予以删除.

SCISSCIAHCI

